 2016,
Vol. 25
2016,
Vol. 25



2. 上海市水产养殖工程技术研究中心, 上海 201306;
3. 水产动物遗传育种中心上海市协同创新中心, 上海 201306
水温对甲壳动物的生长和发育具有重要的作用,诸多学者进行了相关研究。曾朝曙等[1]、王洪全和黎志福[2]与赵云龙等[3-4]分别报道了水温对锯缘青蟹、中华绒螯蟹和红螯螯虾胚胎发育的影响;洪万树和刘昌欣[5]与王战芳等[6]分别进行了斑节对虾幼体和中华锯齿米虾生物学零度及有效积温的研究;王博君[7]初步探索了河蟹早繁育苗中胚胎发育积温和控温的规律;刘吉明[8]分析了梭子蟹人工育苗中的积温与其性腺发育的数量关系;洪美玲等[9]报道了不同温度胁迫方式对中华绒螯蟹免疫化学指标的影响;王玉芬[10]研究了温度对饥饿状态下中华绒螯蟹大眼幼体存活及恢复的影响;温小波等[11]报道了温度对中华绒螯蟹幼蟹饥饿代谢的影响;黄晓荣等[12]报道了16 ℃的条件下,中华绒螯蟹整个胚胎发育过程需40 d 左右,有效积温达到448.3 ℃·d;HARTONLL[13]认为水温、水质和食物是影响甲壳动物生长和蜕壳最重要的环境因素; LEFFLER[14] 、CHUNG等[15]报道了水温的季节性变化直接影响蓝蟹,在寒冷的季节蓝蟹的生长和繁殖停止,而随着温度的升高生长和繁殖又慢慢恢复;另外,SHI等[16]认为在每天水温不低于15 ℃时,罗非鱼的肉增重系数与积温的相关性极为显著。
中华绒螯蟹(Eriocheir sinensis)俗称河蟹,大闸蟹、毛蟹,属于节肢动物门(Arthropoda)、甲壳纲(Crustaces),是我国重要的水产养殖品种之一。尽管国内外学者在水温对甲壳动物生长和发育的影响方面进行了相关研究,但中华绒螯蟹群体发育过程中生长和蜕壳与有效积温的研究方面还未有详细报道。本实验将小规模网箱养殖与生态池塘养殖相结合,在池塘中放置网箱养殖中华绒螯蟹,系统地观察并记录了中华绒螯蟹幼蟹的群体发育过程和水温变化情况,为进一步研究中华绒螯蟹生长、蜕壳与有效积温的关系提供理论依据。
1 材料与方法 1.1 实验地点本实验于2014年5-10月在上海市崇明县新河镇新建村上海福岛水产专业养殖合作社(31°36′N ~ 31°33′N、121°33′E ~ 121°30′E)基地进行。该区地处北半球亚热带,属亚热带湿润季风气候,全年的日照数为2 094.2 h左右,年平均气温15.2 ℃。且环江靠海,雨量充沛,年平均降水量1 025 mm,平均海拔3.7 m,土壤肥沃,适宜河蟹的生长。
1.2 实验材料RC-4温度记录仪:可连续记录温度,可选配外置传感器,并且自动切换到外置传感器;记录仪通过USB连接电脑后,能直接上传数据;测温范围: 内置探头-30 ℃ ~+60 ℃,外置探头-40 ℃~+85 ℃;在-30 ℃~+60 ℃测温范围内,测量精度为±1 ℃,其余±1.5 ℃;分辨率:0.1 ℃;记录容量:16 000组;记录间隔:10 s~24 h可连续设置。
试验蟹种为中华绒螯蟹蟹种,母本和父本体质量分别为(150.4±5.8) g和(300.2±9.6) g,由上海市中华绒螯蟹产业技术体系苗种基地提供。实验池塘长和宽为59 m和38 m,面积2 242 m2,实验网箱为拉链式全封闭网箱,材质尼龙,前期网箱网目大小为1.25 mm;后期网目为4.25 mm,网箱长×宽×高为2.0 m×1.0 m×1.5 m。
1.3 实验设计在同一生态池塘中,放置4口网箱(图 1),每排网箱间距为2 m,每口网箱占地面积2 m2,实验区域位于池塘正中央。将4个RC- 4温度记录仪做好防水后,分别固定在固定网箱的竹竿上;探头放入网箱内入水深度为20 cm,且每周清理一次;每次记录时间间隔为20 min ,静待实验开始时开启,开启后每天至少早晚各查看一次。2014年5月15日,将大眼幼体按每箱500只放养于实验网箱中,大眼幼体均质量为(6.0±0.5) mg。每口网箱投放等量沉水植物伊乐藻(Elodea nuttallii)和浮水植物水花生(Altemanthena philoxeroides),并确保在养殖过程中水草覆盖面积达60%左右。在各网箱中央放置一饵料盘,长×宽为0.3 m×0.5 m,用绳系于网箱顶,可拉出水面投放饵料和观察河蟹的摄食情况。饲料每天17:00按王武等[17]方案投喂(表 1),并根据实际摄食情况适量增减。网箱每4天清理一次,以保证网箱的透水性。

|
图 1 实验网箱分布图
Fig. 1 Test cages distribution
1.伊乐藻区; 2.水花生区; 3.环沟; 4.环沟内区域; 5.塘埂与环沟之间区域。 1. Elodea nuttallii; 2. Altemanthena philoxeroides; 3. Circular groove; 4. The area in circular groove; 5. The area between the pond dam and circular groove. |

|
表 1 蟹种在不同发育阶段的投饵状况和要求 Tab.1 The feeding conditions at different developmental stages and requirements of the Chinese mitten crab |
采样时间:根据河蟹蜕壳周期,在河蟹大批蜕壳(80%~85%)完成后蟹壳刚固化时采样。采样时间根据蟹种蜕壳周期和每天的细致观察得出。采样方式和数量:网捕,随机采样,不分大小,每个网箱≥ 30只。采样共11次,分别对应河蟹的11次蜕壳。测量指标包括:体质量、壳长、壳宽和体高。体质量称量工具和方法:分析天平,单个称量,称量前用吸水纸吸去蟹壳外部水分。壳长、壳宽、体高测量工具和方法:电子数显卡尺,精确度0.01 mm,以头中部凹陷处到尾部垂线距离为壳长;以壳的最宽处作为壳宽;以蟹体背部中央最高处到腹部的垂直距离为体高。
水质对中华绒螯蟹的生长发育非常重要,因而对其进行了全程监测,测量周期为10 d一次。水质指标监测结果:水温为19.8~31.4 ℃,pH为7.3~8.4,溶解氧为5.0~15.0 mg/L,氨氮(NH4+-N)含量小于0.1 mg/L,亚硝酸盐氮小于0.1 mg/L。水质指标均在中华绒螯蟹适宜生长范围之内。水样通过专业的有机玻璃采水器采取,采取水样当场测定溶氧、pH、温度,其他指标全部在2 h内带回实验室测定。溶解氧和水温采用美国YSI550A溶氧仪测定,pH采用PHSJ-3F型实验室pH酸度计测定,氨氮采用纳氏试剂光度法测定,亚硝酸盐(NO2--N)采用盐酸萘乙二胺比色法测定。
1.5 蟹种分类及鉴别根据王武教授蟹种鉴别方法[17]将蟹种分为4类:正常蟹、早熟蟹、蜘蛛蟹和残蟹。
早熟蟹的特征鉴别:(1)性成熟蟹种的雌蟹腹部已成团脐;雄蟹螯足绒毛及步足刚毛稠密,且颜色较深,交接器变成坚硬骨质化的管状体。(2)性成熟的蟹种背部凹凸不平,颜色为墨绿色或青色。(3)性成熟蟹种性腺已经发育,打开蟹种的头胸甲,在肝区上可看到有两条紫色条状物且有卵粒,或有两条白色块状物即精巢,则表明性腺已成熟。
蜘蛛蟹的特征鉴别:蟹种长成收获时,个体大小和体质量远远小于平均体质量的河蟹。
残蟹的特征鉴别:附肢残缺不全或体表有重大伤残的蟹。
1.6 计算方法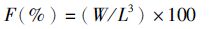 (1)
(1)
式中:F为肥满度; W为体质量;L为体长。
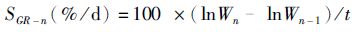 (2)
(2)
式中:SGR-n为第n次蜕壳后的特定生长率;Wn为第n次蜕壳后河蟹体质量;Wn-1为第n-1蜕壳后河蟹体质量;t为养殖天数或蜕壳所需天数。
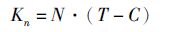 (3)
(3)
式中:Kn为中华绒螯蟹第n次蜕壳所需要的有效积温(℃·d); N为中华绒螯蟹完成第n次蜕壳所需要的时间(d);T为发育期间的平均水温(℃);C为中华绒螯蟹发育的阈温度又称绝对生长零度(℃),根据黄晓蓉等[12]计算为5 ℃。
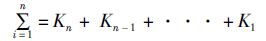 (4)
(4)
式中: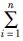

试验数据采用Excel和SPSS 22.0进行作图、统计和方差分析。
2 结果 2.1 池塘水温的变化规律从总体趋势上看,5月至10月水温先升高后下降,其中5月30号至6月6号、6月16号至7月7号、8月11号至8月22号出现幅度较大、时间较长的降温;平均水温为(24.4±0.05)℃;最低和最高温为(17.8±0.04)℃和(31.4±0.08)℃,分别出现在2014的5月16日和8月6日(图 2)。

|
图 2 池塘平均水温变化
Fig. 2 The changes of the average water temperature in the pond
|
从总体趋势上看,蜕壳间期(第一次蜕壳间期从苗种淡化完成后刚放进育苗池塘开始算起)和有效积温均随着蜕壳次数的增加呈逐渐升高趋势(图 3、表 2)。蜕壳间期前4次增长相对缓慢,第5~7次增长较快、第8和第9次变缓、第10和11次又急剧增长,增长趋势较接近指数函数y=2.191 1e0.2568x(R2= 0.980 4),见图 3a。有效积温前4次升高较平稳,第5 ~11次升高较剧烈,增长趋势最接近指数函数y=40.942e0.2655x(R2=0.990 4),见图 3b。累计有效积温增长趋势最接近多项式y=33.336x2-115.05x+196.14(R2= 0.997 8),见图 3c。

|
图 3 中华绒螯蟹11次蜕壳所需的有效积温、
累计有效积温和蜕壳间期增长变化
Fig. 3 The eleven times molting changes in effective ac-
cumulated temperature,cumulative effective accumulated
temperature and molt interval of the Chinese mitten crab
|

|
表 2 中华绒螯蟹一龄蟹11次蜕壳后生长性状数据 Tab.2 The growth traits data at eleven times molt of the Chinese mitten crab |
从总体趋势上看,特定生长率随着河蟹蜕壳呈现先升高后降低的趋势:第1次到第2次升高,第2次之后逐渐降低,第3 ~ 6次降低较快,尤以第4、6次最为明显,分别降低(6.448±0.433)%/d和(5.315±0.417)%/d;第7~11次则下降缓慢,以第8次最为明显,降低(0.332±0.018)%/d。另外,大眼幼体到蟹种的平均特定生长率为(4.510±0.377)% / d(图 4、表 2)。

|
图 4 中华绒螯蟹11次蜕壳的特定生长率变化
Fig. 4 The eleven times molting changes in
specific growth rate of the Chinese mitten crab
|
对中华绒螯蟹11次蜕壳后的特定生长率与有效积温的关系作回归曲线后发现(图 5) 。特定生长率随有效积温的变化曲线最接近幂指数增长关系,公式为y=20.926e-0.005x, R2=0.933 3。

|
图 5 中华绒螯蟹11次蜕壳后特定生长率与
有效积温的关系
Fig. 5 The function of specific growth rate and effective
accumulated temperature after eleven times molting
for the Chinese mitten crab
|
我们对中华绒螯蟹的壳长、壳宽和体高与累计有效积温的关系作回归曲线(图 6、表 2)。壳长、壳宽、体高均随累计有效积温的增长均呈幂指数增长关系,累计有效积温对壳长、壳宽、体高的相关性极为显著。且曲线变化的程度不同说明了随着有效积温的增长,体高变化最为显著,其次为壳长和壳宽。另外,体质量随累计有效积温的增长趋势最接近多项式y=0.000 6x2+0.214 8x-27.208(R2= 0.998 5)(图 7)。

|
图 6 中华绒螯蟹的壳长、壳宽和体高与累计有效积温的关系
Fig. 6 Cumulative effective accumulated temperature
as a function of shell length,shell width and body
height for the Chinese mitten crab
|

|
图 7 中华绒螯蟹的体质量与累计有效积温的关系
Fig. 7 The function of weight and cumulative
effective accumulated temperature of
the Chinese mitten crab
|
从总体趋势上看,肥满度并非是随着有效积温的增长而一直增长的,而是有增有降(图 8、表 2)。在仔蟹阶段(第Ⅰ~Ⅲ期蜕壳)肥满度增长相对较快;第4次蜕壳后,中华绒螯蟹进入幼蟹阶段,第4、5次蜕壳后肥满度增长明显减慢,增幅为0.01%±0、0.02%±0;到第6、7次蜕壳后肥满度增长加快,但到第8次蜕壳后,肥满度没有增长反而下降,降幅达4.42%±0;第9、10次蜕壳后肥满度恢复增长,第10次蜕壳后,肥满度增长率最大,增幅为10.76%±0.02%;但第11次蜕壳后肥满度又降低,降幅达到最大,为6.57%±0.01%。

|
图 8 中华绒螯蟹11次蜕壳后肥满度与
有效积温的关系
Fig. 8 The function of relative fatness and effective
accumulated temperature after eleven times
molting for the Chinese mitten crab
|
第11次蜕壳后,中华绒螯蟹蟹成长为一龄蟹种,平均成活率为28.43%±4.61%,正常蟹平均成活率为22.03%±4.07%。
蟹种收获后,将收获的蟹种以网箱为单位进行分类统计,将每口网箱内蟹种按正常蟹、早熟蟹、蜘蛛蟹、残蟹进行分类统计,并求出各类蟹所占百分比的平均值。结果,百分比最高的是正常蟹(77.50%±2.32%),次之是残蟹(14.21%±1.20%),再者为蜘蛛蟹(4.82%±1.19%),最后为早熟蟹(2.36%±0.41%)。
3 分析与讨论 3.1 水温是中华绒螯蟹生长发育的重要因素水温对水生动物的摄食、生长和繁殖等都有重要影响,每种水生动物都有其适宜生长的水温范围和最适生长水温。一般来说,水生动物在其适宜生长水温范围内,随着温度的升高,其摄食量也逐渐增大,生长速度也逐渐加快。这个范围的水温维持时间越长,水生动物的个体增长越快[18]。中华绒螯蟹也是如此,在低于5 ℃时基本不摄食,且对高温适应能力差,特别是幼蟹,在30 ℃以上水域中的生长时间过长,就容易产生性早熟[17]。
另外,水生动物需要水温达到一定值时才能够生长,这个值在生物学中称为生物学零度,但仅仅水温达到所需尚不足以满足个体生长蜕壳所需的热量条件,还需要一定温度和时间的结合,即需要一定的总热量,称为有效积温。对于中华绒螯蟹有效积温的研究仅胚胎发育时期有相应的报道,王博君[6]认为中华绒螯蟹早繁育苗胚胎发育不仅要有一定的有效积温,在胚胎发育的不同时期对温度也有不同的要求,亲蟹交配产卵期间水温应控制在10 ~ 15 ℃,抱卵蟹越冬和暂养期为6~8 ℃及8 ℃以上,加温促孵期为9~19 ℃,控温时间不少于49 d,这样积温总量在666.7 ℃·d以上,且5年的实践证明采用这个积温控温规律,所繁育的早繁苗在数量、质量上都取得了很好的效果。韦受庆等[19]认为锯缘青蟹在18 ~ 28 ℃温度条件下,从排卵到卵裂需要16 h;薛俊增等[20]认为三疣梭子蟹在11.5~12.0 ℃条件下,从排卵到卵裂需52 h;黄晓蓉等[13]认为与前两种蟹类相比,中华绒螯蟹胚胎发育所需时间较长,16 ℃的水温中发育约需40 d,有效积温达到448.3 ℃·d。而我们监测了中华绒螯蟹胚胎发育之后水温情况,并计算了有效积温,得出幼蟹阶段总有效积温为(3 040.4±244.16) ℃·d,且随着蜕壳次数的增加有效积温增长趋势最接近指数函数y=40.942e0.2655x(R2=0.990 4),累计有效积温增长趋势最接近多项式y=33.336x2-115.05x+196.14(R2=0.997 8)。
3.2 特定生长率、壳长、壳宽等与水温的关系陈立侨等[21]在实验室条件下研究了水温、饵料和投饵率对河蟹生长的影响,认为水温在10~26 ℃范围内,河蟹对不同饵料的摄食率和特定生长率均与水温呈正相关关系。张庆阳等[20]研究了稻田网箱养殖辽河水系中华绒螯蟹幼蟹的个体生长后得出,大眼幼体长成蟹种后雄蟹的特定生长率为(7.176 5%±0.168 4%)/d,雌蟹的特定生长率为(7.283 3%±0.174 3%)/d。辽河水系的雌、雄蟹特定生长率均高于我们研究的长江水系[(4.510±0.377)%/d],原因之一可能是辽河水系中华绒螯蟹因长期的自然选择在遗传上形成了辽河水系中华绒螯蟹群体生长周期短的特点。
黄姝[22]在实验室条件下观察了中华绒螯蟹成蟹的蜕壳与生长后认为,壳长、壳宽与温度有显著的相关性(P< 0.05),体质量与水温的相关性不显著(P>0.05),但相关系数在 0.9 以上,P值介于 0.05~0.10 之间。而我们对中华绒螯蟹的壳长、壳宽和体高与累计有效积温的关系作回归曲线后认为,壳长、壳宽、体高随累计有效积温的增长均呈幂指数增长关系,累计有效积温对壳长、壳宽、体高的相关性极为显著,且体质量随累计有效积温的增长趋势最接近多项式y=0.000 6x2+0.2148x-27.208(R2= 0.9985),另外,我们还得出肥满度并非是随着有效积温的增长而一直增长的,而是有增有降。
3.3 中华绒螯蟹蜕壳与有效积温的规律王武等[17]将中华绒螯蟹的个体发育划分为幼体、幼蟹和成蟹3个阶段。其中,幼蟹阶段指大眼幼体到一龄蟹种。张庆阳等[23]在实验室网箱条件下对辽河水系中华绒螯蟹进行研究发现,其幼蟹阶段蜕壳次数为11次,与本实验对中华绒螯蟹幼蟹阶段的观察研究情况相同,且蜕壳次数均较王武等推测的多3次。
我们研究还发现,中华绒鳌蟹每次蜕壳后体质量、壳长、壳宽和体高数据发生突跃式生长,而未蜕壳的中华绒鳌蟹几乎停止生长,这与杨培根等[24]笼养中华绒螯蟹的研究结果相近。随着幼蟹的生长,有效积温由最初的49.5 ℃·d增长到692.0 ℃·d,增长了近14倍。我们认为有效积温的不断增加,很可能是随着中华绒螯蟹个体生长,其体积不断增加,为下一次蜕壳所要积累的物质和能量也不断增加,所以有效积温不断增加。由于本实验是在池塘网箱中进行的,实验结果也可能会存在一定的偶然因素,今后会进行室内的对比实验,进一步探讨长江水系中华绒螯蟹幼蟹的生长特点。
| [1] | 曾朝曙, 王桂忠, 李少菁.锯缘青蟹胚胎发育的观察及温度影响胚胎发育的研究[J]. 福建水产, 1991: 45–50. ZENG C S, WANG G Z, LI S J.Observation on development of Scylla serrata embryos and influence of temperature on embryonic development[J]. Fujian Fisheries, 1991: 45–50. |
| [2] | 王洪全, 黎志福.水温、盐度双因子交互作用对河蟹胚胎发育的影响[J]. 湖南师范大学自然科学学报, 1996, 19(3): 63–66. WANG H Q, LI Z F.The alternate effects of water temperature and salinity on embryonic development of the Chinese mitten crab Eriocheir sinensis[J]. Journal of Natural Science of Hunan Normal University, 1996, 19(3): 63–66. |
| [3] | 赵云龙, 孟凡丽, 陈立侨, 等.不同水温对红螯螯虾胚胎发育的影响[J]. 湖泊科学, 2000, 12(1): 59–62. DOI:10.18307/2000.0109 ZHAO Y L, MENG F L, CHEN L Q, et al.Effects of different gradient temperatures on embryonic development of Cherax quadricarinatus (Crustacea, Decapoda)[J]. Journal of Lake Sciences, 2000, 12(1): 59–62. DOI:10.18307/2000.0109 |
| [4] | 赵云龙, 堵南山, 赖伟.不同水温对中华绒螯蟹胚胎发育的影响[J]. 动物学研究, 1993, 14(1): 49–53. ZHAO Y L, DU N S, LAI W.Effects of different gradient temperature on embryonic development of the Chinese mitten-handed crab, Eriocheir sinensis (Crustacea, Decapod)[J]. Zoological Research, 1993, 14(1): 49–53. |
| [5] | 洪万树, 刘昌欣.斑节对虾幼体发育的有效积温、生物学零度和耐温实验研究[J]. 福建水产, 1997: 1–6. HONG W S, LIU C X.Study on the effective accumulative temperature, biological zero degree and tolerance of temperature of the larva in the Penaeus monodon[J]. Journal of Fujian Fisheries, 1997: 1–6. |
| [6] | 王战芳, 崔晓东, 巩海雪, 等.中华锯齿米虾生物学零度及有效积温的研究[J]. 河北渔业, 2014: 10–12. WANG Z F, CUI X D, GONG H X, et al.Study on the effective accumulative temperature and biological zero degree of Neocaridina denticulate sinensis[J]. Heibei Fisheries, 2014: 10–12. |
| [7] | 王博君.河蟹早繁育苗胚胎发育积温的调控[J]. 淡水渔业, 1999, 29(8): 45–46. HWANG B J.Regulation on the embryonic development accumulative temperature of crab spawning beforehand[J]. Freshwater Fisheries, 1999, 29(8): 45–46. |
| [8] | 刘吉明.梭子蟹胚胎发育与积温关系初步分析[J]. 河北渔业, 2013: 1–2. LIU J M.Preliminary analysis of the relationship between embryonic development and accumulative temperature of Portunus trituberculatus[J]. Heibei Fisheries, 2013: 1–2. |
| [9] | 洪美玲, 陈立侨, 顾顺樟, 等.不同温度胁迫方式对中华绒螯蟹免疫化学指标的影响[J]. 应用与环境生物学报, 2007, 13(6): 818–822. HONG M L, CHEN L Q, GU S Z, et al.Effect of temperature change on immunochemical indexes of Eriocheir sinensis[J]. Chinese Journal of Applied & Environmental Biology, 2007, 13(6): 818–822. |
| [10] | 王玉芬.温度对饥饿状态下中华绒螯蟹大眼幼体存活及恢复的影响[J]. 河北渔业, 2001: 10. WANG Y F.The effect of temperature on survival and recovery under starvation state of megalopa of the Chinese mitten crab[J]. Heibei Fisheries, 2001: 10. |
| [11] | 温小波, 陈立侨, 艾春香, 等.温度和体重对中华绒螯蟹幼蟹饥饿代谢的影响[J]. 水生生物学报, 2003, 27(4): 357–361. WEN X B, CHEN L Q, AI C X, et al.Effects of temperature and body weight on fasting metabolism of the juvenile Eriocheir sinensis[J]. Acta Hydrobiologica Sinica, 2003, 27(4): 357–361. |
| [12] | 黄晓荣, 庄平, 章龙珍, 等.中华绒螯蟹胚胎发育及几种代谢酶活性的变化[J]. 水产学报, 2011, 35(2): 192–199. HANG X R, ZHUANG Y, ZHANG L Z, et al.Embryonic development and the variation of some metabolism enzyme activity during embryonic development of Chinese mitten crab (Eriocheir sinensis)[J]. Journal of Fisheries of China, 2011, 35(2): 192–199. |
| [13] | HARTNOLL R G.The Biology of Crustacea[M].New York: Academic Press, 1982: 111-185l. |
| [14] | LEFFLER C W.Some effects of temperature on the growth and metabolic rate of juvenile blue crabs, Callinectes sapidus, in the laboratory[J]. Marine Biology, 1972, 14(2): 104–110. DOI:10.1007/BF00373209 |
| [15] | CHUNG J S, PITULA J S, SCHOTT E, et al.Elevated water temperature increases the levels of reo-like virus and selected innate immunity genes in hemocytes and hepatopancreas of adult female blue crab, Callinectes sapidus[J]. Fish & Shellfish Immunology, 2015, 47(1): 511–520. |
| [16] | SHI W L, WEI T M, LI J Q, et al.Effects of accumulated temperature and stocking size on Tilapia nilotica culture in net cages[J]. Shandong Fisheries, 1997, 4(4): 33–35. |
| [17] | 王武, 王成辉, 马旭洲.河蟹生态养殖[M].北京: 中国农业出版社, 2013: 20-239. WANG W, WANG C H, MA X Z.The ecological culture of Eriocheir sinensis[M].Beijing: China Agriculture Press, 2013: 20-239. |
| [18] | 陈万光.几种环境因子对水生动物的影响研究[J]. 洛阳师范学院学报, 2002, 21(5): 133–135. CHEN W G.Research on several environmental factors' influences on aquatic animals[J]. Journal of Luoyang Teachers College, 2002, 21(5): 133–135. |
| [19] | 韦受庆, 罗远裕.青蟹胚胎发生的研究[J]. 热带海洋, 1986, 5(3): 57–62. WEI S Q, LUO Y Y.A study of the embryonic development of Scylla serrata (Forskal)[J]. Tropic Oceanology, 1986, 5(3): 57–62. |
| [20] | 薛俊增, 堵南山, 赖伟.三疣梭子蟹胚胎发育过程中卵内幼体形态[J]. 动物学报, 2001, 47(4): 447–452. XUE J Z, DU N S, LAI W.Morphology of egg-larvae of swimming crab (Portunus trituberculatus) during embryonic development[J]. Acta Zoologica Sinica, 2001, 47(4): 447–452. |
| [21] | 陈立侨, 堵南山, 赖伟.水温 饵料 投饵率对河蟹生长和饵料转化率的影响[J]. 水产科技情报, 1993, 20(4): 153–157. CHEN L Q, DU N S, LAI W.Effects of water temperature, feeds and ration size on the growth and the feed conversion ratio of Chinese mitten crab (Eriocheir sinensis)[J]. Fisheries Science & Technology Information, 1993, 20(4): 153–157. |
| [22] | 黄姝. 实验室条件下中华绒螯蟹成蟹的蜕壳、生长观察与蜕皮激素受体基因的克隆、表达分析[D]. 上海: 上海海洋大学, 2014: 4-10. HUANG S. Observation on molting and growth, molecular cloning and expression of ecdysone receptor (EcR) gene in adult Chinese mitten crab in the laboratory condition[D]. Shanghai: Shanghai Ocean University, 2014: 4-10. http://cdmd.cnki.com.cn/article/cdmd-10264-1014051740.htm |
| [23] | 张庆阳, 马旭洲, 王昂, 等.稻田网箱养殖辽河水系中华绒螯蟹幼蟹的个体生长[J]. 动物学杂志, 2015, 50(1): 112–121. ZHANG Q Y, MA X Z, WANG A, et al.The research of individual growth of Eriocheir sinensis for Liaohe population juvenile crab in paddy field net cage[J]. Chinese Journal of Zoology, 2015, 50(1): 112–121. |
| [24] | 杨培根, 李晨虹.笼养中华绒螯蟹的蜕壳与生长[J]. 上海水产大学学报, 1998, 7(2): 158–161. YANG P G, LI C H.Molting and growth of Chinese mitten crab, Eriocheir sinensis, raised in plastics cage[J]. Journal of Shanghai Fisheries University, 1998, 7(2): 158–161. |
2. Shanghai Engineering Research Center of Aquaculture, Shanghai 201306;
3. Shanghai Collaborative Innovation Center for Aquatic Animal Genetics and Breeding, Shanghai 201306, China