 2016,
Vol. 25
2016,
Vol. 25



2. 三门县东航水产育苗科技有限公司, 浙江 三门 317100
缢蛏(Sinonovacula constricta)俗称蛏子、青子,隶属于软体动物门(Mollusca),瓣鳃纲(Lamellibranchia),异齿亚纲(Heterodonta),帘蛤目(Veneroida),竹蛏科(Solenidae),缢蛏属(Sinonovacua),是一种埋栖型的双壳经济贝类,广泛分布于中国、日本和朝鲜等国的沿海地区,是我国四大海产品贝类之一[1]。我国在20世纪90年代初开展了缢蛏的人工育苗,并且取得成功,目前养殖单位主要采用人工苗进行养殖生产,且用于繁殖人工苗的亲贝多数也来自人工养殖的缢蛏,随着多年的人工养殖、频繁的近交必定会导致物种多样性的下降,造成缢蛏种质资源的退化[2]。其中由于抗逆性下降导致缢蛏存活率低、抗病力弱等问题显得尤为突出。
盐度和温度都是影响海洋无脊椎动物生长发育的重要环境因素[5-7]。盐度的变化会直接导致海洋生物渗透压的失衡,进而影响海洋生物的能量收支、呼吸排泄[3]。文蛤幼体在适宜的盐度范围内,其成活率、变态率和生长速度都最高[4]。泥蚶在不同的发育阶段,其最适生存盐度、最适生长盐度不尽相同[5]。有研究表明,温度越高,栉孔扇贝的死亡越严重,温度与病害暴发有直接的相关性[8]。在一定的温度范围,多数贝类的代谢率会随着温度的升高而升高,当超过其承受最高温范围后,会引起贝类生理功能的紊乱[8-9],最终导致贝类的死亡[10-11]。
缢蛏是一种典型的广温、广盐性海洋贝类,但温度或盐度过高都会引起缢蛏相应的应激反应,导致其成活率、抗病力下降,最终受病害感染而死亡。林笔水等[12-14]采用单因子和多因子结合的方法,研究了温度和盐度对缢蛏幼体存活、生长及发育的影响。范德朋等[15]研究了温度对缢蛏耗氧率和排氨率的影响。吕昊泽等[16]的研究表明盐度能影响缢蛏超氧化物歧化酶和过氧化氢酶的活性,进而调节自身对环境的适应性。我国缢蛏养殖北自辽宁沿海南至广东、广西,适合河口半咸水海域生长的缢蛏常常会遇到高温和高盐海水的影响,抗逆性能不高时常会出现成活率低、生长缓慢、个体养不大等生产性难题,及时开展缢蛏耐高盐、耐温等抗逆性状的选育将利于缢蛏养殖业在我国各地的持续发展。但到目前为止,关于利用盐度和温度对不同缢蛏家系的影响,来达到缢蛏抗逆性家系选育的报道尚未见到。家系选择是进行新品种选育的最直接、最有效的方法之一[17]。国内外对很多经济贝类如马氏珠母贝(Pinctada martensii)[18]、海湾扇贝(Argopenten irradians)[19]、牡蛎(Ostrea edulis,O.chilensis,Crassostrea virginica)[20]、文蛤(Meretrix meretrix)[21]等都开展了选育且取得了较好的进展,我国已获得了不少贝类新品种,如中国科学院海洋研究所的文蛤“科浙1号”、厦门大学的杂色鲍“东优1号”等。本研究是在乐清湾的优良群体子四代的基础上,于2014年建立并获得了生长性状差异的家系,对不同缢蛏家系耐热、耐高盐性能方差分析和多重比较,并对生长和耐热、耐高盐性能作相关分析,旨在为缢蛏的生长和抗逆性状选育工作提供依据。
1 材料与方法 1.1 实验材料本实验缢蛏家系的建立在浙江省三门县六敖镇东航水产育苗科技有限公司进行,2014年基于乐清湾的优良群体子四代,采取单配对的方法建立并获得了27个全同胞家系,同时还得到了缢蛏群体繁育材料,经过3个月的室内培育和对各家系生长数据定期的测量,获得了5个具有生长优势的家系,17个平均生长性能和5个生长劣势的家系。之后将各家系和群体繁育材料放在用渔网分隔好的室外土池暂养,再经过3个月室外培育到幼贝后,将5个生长优势、生长劣势的家系和缢蛏群体繁育材料挖出,分别作为实验用家系组和对照组。
1.2 各家系幼蛏室内暂养实验塑料桶(100 L)底部铺5 cm厚软泥土,水位高出土面60 cm,将土池中挖出来的10个家系和群体繁育幼蛏分别放在11个试验桶中暂养,每桶放幼蛏300粒左右,试验期间,持续微充气,投喂球等鞭金藻 (Isochrysis.galbana)、牟氏角毛藻(Chaeroeeros moelleri)、亚心型扁藻(Platymonas subcordiformis)的混合藻液。保证水中饵料充足,各组每天换水1/2,塑料桶放置在室内,自然光照,水温25 ℃,盐度13,pH 7.8。
1.3 各家系幼蛏生长性能测定对各实验家系和群体繁育幼蛏进行生长测量。测量指标主要包括壳长、总重和壳长日增长率,其中壳长日增长率是指幼蛏从土池挖出来后,室内培育条件下的壳长日增长率。日增长率=(结束时壳长-开始时壳长)/ 测量天数,每个家系用相同的方法重复测量3次,每个重复随机测定30粒幼蛏,计算各家系幼蛏壳长、总重和壳长日增长率的平均数,评价不同家系的生长性能。壳长测量采用数显游标卡尺,总重测量时用滤纸吸干体表水分,用SHIMADZUAUY 120(精确至0.1 mg)电子天平测量。
1.4 方法 1.4.1 半致死温度、盐度的确定各家系幼蛏经过一个星期室内暂养后,从每个家系随机挑选出45粒幼蛏混养,再分别养殖于15个实验用塑料桶中,塑料桶培养条件同1.2,每桶放30粒幼蛏。根据缢蛏参考文献和其他贝类温度实验,实验水温设置在30~38 ℃,以2 ℃为梯度,预实验设置5个家系组和一个常温群体繁育空白对照组,每组按照相同实验方法重复3次。温度控制采用曹伏君等[22]方法,使用水银温度计、500 W石英加热管、1 000 W电子继电器及人工冰袋控温,温度精确控制在±0.2 ℃。以幼蛏50%死亡的温度为生存临界温度,即半致死温度,以平均相对存活率为依据,采用二点法[23],计算高温端生存临界温度(LS50)。公式为:
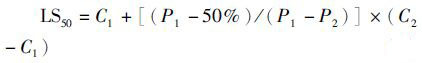 (1)
(1)
式中:C1、C2分别为存活率接近50%的高端温度(℃);P1、P2为相应的相对存活率。死亡的界定以贝壳张开、外套膜收缩为标准。
半致死盐度的确定与温度预实验类似,盐度设置为30~38,以2为盐度梯度,设置5个家系组和一个常盐群体繁育空白对照组,每组按照相同实验方法重复3次。盐度升高是通过向过滤海水中加海盐精实现,半致死盐度同样采用二点法获得。
1.4.2 家系耐热、耐高盐性实验耐热性实验是在室内塑料桶中进行,幼蛏按30粒/家系分别放入10个实验塑料桶中,培养的初始条件同1.2。实验以常温常盐下的群体繁育组幼蛏作空白对照,每组按照相同实验方法重复3次。盐度13保持不变,按每4小时升温1 ℃的速率达到半致死水温后统计数据,实验期间,持续微充气加藻,加藻前需消除藻液的温差,以免影响各家系组温度。每天等温换水1/2。统计幼蛏死亡率直至实验结束。
耐高盐性实验条件与方法与耐热性实验相似,保持水温为25 ℃,盐度按每小时升高2的速率调到半致死盐度后统计,实验期间,持续微充气加藻,加藻前需消除藻液的比重差,以免影响各家系组盐度。每天等盐度换水1/2。统计幼蛏死亡率直至实验结束。
1.5 指标测定存活率和相对存活率的测定
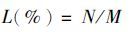 (2)
(2)
式中:L是存活率;N是实验组中幼蛏存活数;M是实验组中幼蛏个体总数。
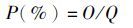 (3)
(3)
式中:P是相对存活率;O是实际观察存活率;Q是对照组存活率。
1.6 数据处理和统计分析利用SPSS 17.0软件对各家系存活率进行单因素方差分析,Turkey HSD对各家系多重比较,并对在高温、高盐条件下各家系幼蛏生长指标与存活率进行相关性分析。
2 结果与分析 2.1 不同家系生长性状分析描述在对27个全同胞家系前期3个月的生长性能评定中,有生长优势的家系为5个,家系编号分别为2号、7号、9号、13号和19号,生长劣势的家系分别为5号、6号、14号、25号和27号家系。经过三个月的室外土池培育后,测定了各家系的平均壳长、总重和壳长日增长率。缢蛏不同家系后期的生长性状统计描述见表 1,前期具有生长优势的5个家系,在平均壳长、总重和壳长日增长率上都显著高于生长劣势家系(P< 0.05)。其中9号家系各项生长指标均值均为最大。27号家系各项生长指标均值均最小,9号家系的壳长、总重和日增长率均值相比27号家系分别高了29.66%、40.00%和32.37%。

|
表 1 不同家系幼蛏的壳长、总重和壳长日增长率 Tab.1 Comparison of growth performance of different families of Sinonovacula constricta |
各采用5个梯度实验对幼蛏半致死温度、盐度进行确定,观察记录并及时捞出死蛏,在实验14 d后幼蛏的死亡情况基本稳定,即以14 d作为家系耐热、耐高盐性能的考察时间,计算存活率和相对存活率。预试验结果表明,随着温度、盐度的升高,幼蛏的存活率呈现下降趋势,在温度为36和38 ℃时,幼蛏的平均相对存活率分别为58.4%和24.8%,采用二点法计算得到幼蛏14 d的半致死温度为36.5 ℃;在盐度为34和36时,幼蛏的平均相对存活率分别为62.6%和48.6%,采用二点法计算得到幼蛏14 d的半致死盐度为35.8。

|
表 2 不同温度、盐度梯度下幼蛏的存活率和相对存活率 Tab.2 Survival rate,relative survival rate of Sinonovacula constricta under different temperature and salinity gradients |
按照上述确定的半致死温度、盐度进行不同幼蛏家系的抗逆性实验,以14 d作为实验的考察时间。结果表明,在半致死条件下,幼蛏在24 h后开始出现水管吐水频率下降、双壳收缩无力等活力下降的症状,3 d后开始出现少量的死亡,死亡界定为水管无收缩、双壳打开,7 d死亡率达到最大值;10 d后死亡趋于稳定。各家系半致死温度下的相对存活率为(41.1%±0.50%) ~(71.7%±0.50%),且在耐热性能上存在着明显差异,5个生长优势家系在生存临界温度下的相对存活率都在66%以上,其中9号家系存活率最高,显著大于其他家系(P< 0.05),有4个家系(5号、6号、25号和27号)存活率在50%以下。
各家系半致死盐度下的相对存活率为(47.7%±0.40%)~(71.5%±0.36%),且在耐高盐性能上存在明显差异,5个生长优势家系在半致死盐度下的相对存活率都在67%以上,其中19号家系在半致死盐度下相对存活率最高,显著大于其他家系(P< 0.05),有3个家系(5号、25号和27号)存活率在50%以下。
本文将相对成活率在70%以上的家系定义为耐热或耐高盐性能强的家系,分别将相对成活率在60%~70%、50%~60%的家系定义为耐热或耐高盐性能较强和一般的家系,相对成活率在50%以下的家系定义为耐热或耐高盐性能差的家系。由此得到3个耐热性能强的家系(7号、9号和13号),2个耐热性能较强的家系(2号和19号),1个耐热性能一般的家系(14号),4个耐热性能差的家系(5号、6号、25号和27号);1个耐高盐性能强的家系(19号),4个耐高盐性能较强的家系(2号、7号、9号和13号),2个耐高盐性能一般的家系(6号和14号),3个耐高盐性能差的家系(5号、25号和27号),见表 3。

|
表 3 各家系在生存临界温度、盐度下的存活率及相对存活率 Tab.3 Survival rate,relative survival rate of different families of Sinonovacula constricta under critical temperature and salinity |
在进行幼蛏家系的耐热、耐高盐性实验前,对各家系的生长指标进行了测量,本文选取各家系的平均壳长与半致死条件下幼蛏的存活率进行Pearson相关性分析。壳长与半致死温度、盐度存活率均为正相关,且相关性显著(P< 0.05)。相关系数分别是0.970和0.920。
3 讨论缢蛏作为一种典型的滩涂贝类,因其营埋栖生活而活动范围有限,当外界温度、盐度环境剧烈变化时,其生存就会受到严重考验。尤其是在夏季退潮后,滩涂存有少量海水,或小潮期在中午前后枯潮,水位较低,潮水经太阳暴晒后水温会急剧上升,高温的海水会从出入水孔传至蛏穴,造成缢蛏的烫伤或烫死,同时连续的干旱会造成海水盐度升高也会引起缢蛏大量死亡[2]。有研究表明,盐度、温度的波动会影响缢蛏体内水分和粗蛋白含量、代谢能力和消化酶活性[15-16],甚至能影响贝类体内免疫应答水平,造成其抗逆能力的下降。因此,从种质水平上强化缢蛏的抗逆性能,才能提高其在高温、高盐环境中的存活率。
温度和盐度是海洋环境中特别重要的两个因素,它们与海洋生物的生长发育有着密切的关系,因其比较容易调节和控制而引起生态学家的广泛关注。很多学者认为这两个因素对生物的影响存在一定的联系。其中CASTAGNA[24]就认为在一定的温度范围内,生物对盐度的反应速率主要是受温度高低的影响,但生物对盐度的耐受限度并不会因温度改变。CALABRESE[25]认为只有当温度和盐度或其中之一接近生物耐受极限范围时,温度和盐度相结合的相互作用才会明显显示出来,相反,当温度或盐度其中之一在安全范围时,则没有明显的关系。在进行耐热、耐高盐性实验时,本实验相应的盐度、温度都控制在最适宜缢蛏生存和生长的范围内,温度或盐度单一因子的实验也能准确反映缢蛏不同家系耐热、耐高盐性能的差异。
林笔水等[13-14]的研究表明,缢蛏稚贝的适温范围为10~35 ℃,适盐范围为4.5~28.3,不同发育时期的缢蛏对水温、盐度的耐受力也不尽相同。我国渤海湾、黄海、东海和南海海域全年平均盐度分别为29.84、30.04、32.37和33.01,四大海域盐度相差不大,但缢蛏多生活在近海河口半咸水海域,再加上夏天多雨,沿海不同地区盐度相差较大。生活在高盐区和低盐区蛏苗的适盐范围相差不大,但高盐区蛏苗比低盐区蛏苗更耐高盐[12];四大海域全年平均温度分别是13.1、16.1、22.4和27.2 ℃,不同盐度海区蛏苗对温度耐受程度亦有不同[12]。本实验得到缢蛏生存临界温度、盐度与林笔水等研究略有差别,可能与本实验缢蛏生活海区的水环境有关。不同生长性能的家系在耐热、耐高盐性能上均表现出差异性,半致死温度和盐度条件下,5个生长优势家系存活率均高于生长劣势家系,耐热、耐高盐性能最强的家系比最差的家系分别高了74%和50%。这与王庆志等[26]关于虾夷扇贝耐高温家系的研究相似。THIVAKARAN等[26]的研究结果表明,螺类对温度和盐度的耐受力与个体大小没有太大的关系,在不同的温度或盐度下,所有大小的个体都能以相似的方式做出反应。这与本实验得到缢蛏对温度、盐度的抗逆性与个体大小有显著性相关的结果不符,本文认为不同家系抗逆性能力的不同,是受到内部因素和外界因素的双重影响,内部因素包括物种之间的差异性、同一物种家系亲本的基因型和遗传力不同等,外界因素包括养殖水环境和个体发育程度等,另外有研究表明在相同养殖时间、养殖条件下,大蛏相对于小蛏对阴干的忍耐性要强[12],本实验保证了各缢蛏家系的外界因素一致,减少养殖条件不同对实验造成的误差,由此,认为这种家系之间抗逆性差异主要由其内部因素的不同造成的。
本研究首次在不同生长性能的缢蛏家系间,开展了其耐热、耐高盐性能的筛选研究,采用家系选育技术,获得了3个耐高温和1个耐高盐家系。为相关基因的筛选提供了重要育种材料,也为如何获得生长性状优良且抗逆性强的缢蛏品种提供了理论依据。
| [1] | 徐凤山, 张素萍.中国海产双壳贝图志[M].北京: 科学出版社, 2008: 211-213. XU F S, ZHANG S P.Chinese marine bivalve illustrated record[M].Beijing: Science Publishing House, 2008: 211-213. |
| [2] | 王兴强, 曹梅, 阎斌伦.缢蛏养殖期间发病原因及防治对策[J]. 现代渔业信息, 2006: 13–16. WANG X Q, CAO M, YAN B L.Analysis of diseases cause and prevention measures in farming of Sinonovacula constricta(Lamarck)[J]. Modern Fisheries Information, 2006: 13–16. |
| [3] | 阎希柱, 王桂忠, 李少菁.盐度对九孔鲍能量收支的影响[J]. 生态学杂志, 2009, 28(8): 1520–1524. YAN X Z, WANG G Z, LI S J.The effects of different salinities on the energy budgets of Haliotis diversicolor aquatilis[J]. Chinese Journal of Ecology, 2009, 28(8): 1520–1524. |
| [4] | 陈冲, 王志松, 隋锡林.盐度对文蛤孵化及幼体存活的生长的影响[J]. 海洋科学, 1999: 16–18. CHEN C, WANG Z S, SUI X L.Effect of salinity on incubation and growth and survival of Meretrix meretrix spat[J]. Marine Sciences, 1999: 16–18. |
| [5] | 尤仲杰, 徐善良, 边平江, 等.海水温度和盐度对泥蚶幼虫和稚贝生长及存活的影响[J]. 海洋学报, 2001, 23(6): 108–113. YOU Z J, XU S L, BIAN P J, et al.The effects of sea water temperature and salinity on the growth and survival of Tegillarca granosa larvae and juveniles[J]. Acta Oceanologica Sinica, 2001, 23(6): 108–113. |
| [6] | 栗志民, 刘志刚, 姚茹, 等.温度和盐度对皱肋文蛤幼贝存活与生长的影响[J]. 生态学报, 2010, 30(13): 3406–3413. LI Z M, LIU Z G, YAO R, et al.Effect of temperature and salinity on the survival and growth of Meretrix lyrata juveniles[J]. Acta Ecologica Sinica, 2010, 30(13): 3406–3413. |
| [7] | 王丹丽, 徐善良, 尤仲杰, 等.温度和盐度对青蛤孵化及幼虫、稚贝存活与生长变态的影响[J]. 水生生物学报, 2005, 5(5): 495–501. WANG D L, XU S L, YOU Z J, et al.The effects of temperature and salinity on the incubation of Cyclina sinensis and survival, growth and metamorphosis of C.sinensis larvae and juveniles[J]. Acta Hydrobiologica Sinica, 2005, 5(5): 495–501. |
| [8] | 杨红生.温度对墨西哥湾扇贝耗氧率及排泄率的影响[J]. 海洋学报, 1998, 20(4): 91–96. YANG H S.Effects of temperature on oxygen consumption and ammonia-N excretion of the southern bay scallop Argopecten irradians concentricus[J]. Acta Oceanologica Sinica, 1998, 20(4): 91–96. |
| [9] | 王俊, 姜祖辉.栉孔扇贝耗氧率和排氨率的研究[J]. 应用生态学报, 2002, 13(9): 1157–1160. WANG J, JIANG Z H.Oxygen consumption and ammonia-N excretion rates of Chlamys farreri[J]. Chinese Journal of Applied Ecology, 2002, 13(9): 1157–1160. |
| [10] | 林君卓, 许振祖.温度和盐度对文蛤幼体生长发育的影响[J]. 福建水产, 1997: 27–33. LIN J Z, XU Z Z.The effects of temperature and salinity on the development of Meretrix meretrix larvae[J]. Journal of Fujian Fisheries, 1997: 27–33. |
| [11] | 刘贤德, 闫玉华, 王志勇, 等.杂色鲍对高温和低盐耐受力的初步研究[J]. 集美大学学报(自然科学版), 2008, 13(4): 301–303. LIU X D, YAN Y H, WANG Z Y, et al.A preliminary study on tolerance to high temperature and low salinity of Haliotis diversicolor reeve[J]. Journal of Jimei University(Natural Science), 2008, 13(4): 301–303. |
| [12] | 吴天明, 林笔水.几种环境因子对缢蛏苗存活率的影响[J]. 台湾海峡, 1987: 2. WU T M, LIN B S.Effects of several environmental factors on survival rate of young shells of Sinonovacula constricta(Lamarck)[J]. Journal of Oceanography In Taiwan Strait, 1987: 2. |
| [13] | 林笔水, 吴天明.温度和盐度同缢蛏稚贝存活及生长的关系[J]. 水产学报, 1986, 10(1): 40–50. LIN B S, WU T M.The relations of temperature and salinity to the survival and growth of the spat of Sinonovacula constricta(Lamarck)[J]. Journal of Fisheries of China, 1986, 10(1): 40–50. |
| [14] | 林笔水, 吴天明.温度与盐度和缢蛏幼体生存、生长及发育的关系[J]. 水产学报, 1990, 14(3): 171–178. LIN B S, WU T M.Temperature and salinity in relating to the survinal growth and development of the larvae and spat of Sinonovacula constricta(Lamarck)[J]. Journal of Fisheries of China, 1990, 14(3): 171–178. |
| [15] | 范德朋, 潘鲁青, 马甡, 等.温度对缢蛏(Sinonovacula constricta)耗氧率和排氨率的影响[J]. 青岛海洋大学学报(自然科学版), 2002, 32(1): 56–62. FAN D P, PAN L Q, MA S, et al.Effects of temperature on oxygen consumption rate and ammonia excretion rate of Sinonovacula constricta[J]. Journal of Ocean University of Qingdao(Natural Science), 2002, 32(1): 56–62. |
| [16] | 吕昊泽, 刘健, 陈锦辉, 等.盐度对缢蛏超氧化物歧化酶和过氧化氢酶活性的影响[J]. 海洋渔业, 2013, 35(4): 474–478. LV H Z, LIU J, CHEN J H, et al.Effects of salinity on activities of SOD and CAT in hepatopancreas of Sinonovacula constricta[J]. Marine Fisheries, 2013, 35(4): 474–478. |
| [17] | 何玉英, 李健, 刘萍, 等.中国对虾家系幼体对氨氮和pH值的耐受性比较[J]. 中国海洋大学学报(自然科学版), 2008, 38(5): 761–765. HE Y Y, LI J, LIU P, et al.Comparison of the resistance to pH value and ammonia in Chinese Shrimp(Fenneropenaeus chinensis) families[J]. Periodical of Ocean University of China(Natural Science), 2008, 38(5): 761–765. |
| [18] | WADA K T, WADA K T.Genetic selection for shell traits in the Japanese pearl oyster, Pinctada fucata martensii[J]. Aquaculture, 1986, 57(86): 171–176. |
| [19] | ZHENG H, ZHANG G, LIU X, et al.Sustained response to selection in an introduced population of the hermaphroditic bay scallop Argopecten irradians irradians Lamarck (1819)[J]. Aquaculture, 2006, 255(1/4): 579–585. |
| [20] | DAVIS C V.Esitmation of narrow-sense heritability for larval and juvenile growth traits in selected and unselected sub-lines of eastern oysters, Crassostrea viginico[J]. Journal of Shellfish Research, 2000, 19(1): 613. |
| [21] | WANG X H, CHAI X L, LIU B Z.Estimation of genetic parameters for growth traits in cultured clam Meretrix meretrix (Bivalvia:Veneridae) using the Bayesian method based on Gibbs sampling[J]. Aquaculture Research, 2011, 42: 240–247. |
| [22] | 曹伏君, 刘志刚, 罗正杰.海水盐度、温度对文蛤稚贝生长及存活的影响[J]. 应用生态学报, 2009, 20(10): 2545–2550. CAO F J, LIU Z G, LUO Z J.Effects of seawater temperature and salinity on the growth and survival of juvenile Meretrix meretrix Linnaeus[J]. Chinese Journal of Applied Ecology, 2009, 20(10): 2545–2550. |
| [23] | 刘志刚, 王辉, 栗志民, 等.墨西哥湾扇贝高起始致死温度的研究[J]. 中国水产科学, 2007, 14(5): 778–785. LIU Z G, WANG H, LI Z M, et al.Upper incipient lethal temperature of Argopecten irradians concentricus Say[J]. Journal of Fishery Sciences of China, 2007, 14(5): 778–785. |
| [24] | CASTAGNS M.Culture of the bay scallop, Argopecten irradians in Virginia[J]. Marine Fisheries Research, 1975, 37: 19–24. |
| [25] | CALBRESE A.Individual and combined effects of salinity and temperature on embryos and larvae of the coot clam, Mulinia lateris[J]. Marine Biology, 1969, 137: 417–428. |
| [26] | THIVAKARAN G A, KASINATHAN R.Salintry, temperature and desiccation tolerance of intertide Gastropoda[J]. Indian Journal of MarineSciences, 1990, 19: 57–60. |
2. Sanmen Eastern Fisheries Limited, Sanmen 317100, Zhejiang, China