 2016, Vol. 25
2016, Vol. 25


杭州湾海域位于北亚热带,是典型的河口海湾。杭州湾营养物质丰富,水质肥沃,饵料生物繁盛,是多种海洋生物产卵、索饵和育肥的场所[1]。杭州湾东北部地处杭州湾门口处,受长江口冲淡水与东海外海水共同影响[2],因此该片海域呈现与典型性河口区不同的生态特征,渔业生物多样性和数量分布也具备特殊性。
近年来,关于杭州湾海洋生物多样性的报道已开展较多。蔡燕红[3]、王晓波等[4]分别研究了杭州湾浮游植物和浮游动物多样性,为全面了解该海域饵料生物的现状提供较为详实的基础资料。张冬融等[5]研究了杭州湾南岸浮游动物分布及与环境因子的关系,得出温度、盐度对浮游动物分布具有显著影响。周永东等[6]、陈渊戈[7]和闫欣等[8]分别对杭州湾北岸仔稚鱼的种类组成、群聚特征和多样性进行研究,提供了该片海域仔稚鱼基础数据,同时提出杭州湾海域渔业资源保护的科学性和合理性。谢旭等[9]介绍了杭州湾河口区鱼类种类组成与数量分布,并得出鱼类优势种已经小型化、低值化的结论。张海波[10]阐述了杭州湾海洋生物保护的重要性,对杭州湾海域生态系统健康状况进行评估。王淼等[11]对杭州湾北部海域张网渔业资源种类组成进行分析,得出优势种为鲚属鱼类和中国毛虾。以上研究主要针对特定海洋生物群体,如饵料生物、仔稚鱼和鱼类开展调查,没有针对渔业资源的全面调查研究。
本研究以杭州湾海域渔业资源多样性和数量分布为研究对象,分析不同时期渔业资源的多样性差异和数量变动规律,探讨了数量分布与环境因子之间的关系及多样性变动规律,以期为杭州湾海域渔业资源调查评估提供基础数据,也为该海域渔业资源保护和生态修复方案的制定等提供基础资料。
1 材料与方法 1.1 采样区域与调查方法分别于丰水期和枯水期在杭州湾东北部海域设置10个站位进行渔业资源调查(图1)。采样用单船底拖网船(功率88.2 kW),网具为单囊有翼网,网口周长30 m,网衣总长25 m,总高2.4 m,扫海面积宽度为10 m,囊网目20 mm。起网后参照《海洋水产资源调查手册》[12],对所有渔获物进行种类鉴定、重量及数量统计,并对所有种类进行生物学测定,包括体长、体重、性比等数据。

|
图1 调查站位 Fig.1 Sampling stations |
物种相似性分析采用Jaccard相似性系数[13],计算丰、枯水期渔业生物物种组成的相似性,计算公式为:
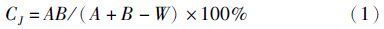
渔业生物资源密度的估算采用扫海面积法:
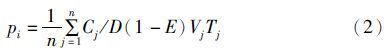
采用Pianka的相对重要性指数[15](Index of Relative Importance,IRI)来衡量物种的优势度:
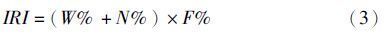
群落多样性分析,采用Shannon-wiener多样性指数(H′),在渔业生物生态研究中,重量多样性与尾数多样性具有不同的生物学含义[19],因此本研究不同站位的渔业生物多样性指数(H′)分别采用重量密度(kg/km2)和尾数密度(103 ind/ km2)计算,公式如下:

利用曲线回归估计(Curve Estimation)分析H′与资源密度之间的相关性。用独立样本t检验分析不同时期平均温度和盐度的显著性差异,显著性水平设为0.05,以上分析方法和检验由SPSS 19.0完成。
2 结果与分析 2.1 种类组成及相似性分别于丰水期和枯水期对杭州湾东北部水域进行渔业资源拖网调查,共捕获40种渔业生物(表1)。其中,丰水期捕获34种,枯水期捕获22种,两时期共有种16种。物种相似性分析表明,两时期的Jaccard相似性系数仅为40%,为中等不相似水平。可见,丰、枯水期杭州湾东北部水域物种相似性水平较低。

|
表1 丰、枯水期杭州湾东北部渔业资源种类组成 Tab. 1 Composition of fishery resource in Northeast of Hangzhou Bay during drought and flood seasons |
丰、枯水期杭州湾东北部渔业资源的平均重量密度分别为294.82 kg/km2和85.76 kg/km2,平均尾数密度分别为83.19×103 ind/km2和33.88×103 ind/km2。
2.2.2 丰水期生物密度的空间分布丰水期渔业资源重量密度最高的站位是6号,为1 402.32 kg/km2(图2a),主要品种为鮸鱼和棘头梅童鱼。此外,8号站位也有较高的重量密度(图2a),为634.72 kg/km2,主要由棘头梅童鱼、白姑鱼和日本蟳构成。

|
图2
丰水期杭州湾东北部渔业资源密度分布
Fig.2
Density distribution of fishery resources in Northeast of Hangzhou Bay in flood season
a为重量密度分布(kg/km2),b为尾数密度分布(103 ind/km2),图3-5同。 a means distribution of weight density(kg/km2),b means distribution of individual density(103 ind/km2),the same in Fig.3-5. |
丰水期渔业资源尾数密度最高的站位是2号,为263.00×103 ind/km2(图2b),主要品种为安氏白虾。此外,6、7和8号站位也具有较高的尾数密度(图2b),分别为122.51×103、172.08×103和122.77×103 ind/km2,主要是因为出现大量安氏白虾的缘故。
2.2.3 枯水期生物密度的空间分布枯水期渔业资源重量密度最高的2个站位是1号和2号(图3a),重量密度分别为152.09 kg/km2和156.14 kg/km2,主要品种为刀鲚和葛氏长臂虾。此外,6、10号站位也具有较高的重量密度(图3a),分别为124.43和120.66 kg/km2,6号站位主要由焦氏舌鳎和髭缟虾虎鱼构成,10号站位主要由半滑舌鳎、髭缟虾虎鱼和葛氏长臂虾构成。

|
图3 枯水期杭州湾东北部渔业资源密度分布 Fig.3 Density distribution of fishery resources in Northeast of Hangzhou Bay in drought season |
枯水期渔业资源尾数密度最高的站位是10号(图3b),为105.14×103 ind/km2,主要品种为葛氏长臂虾。其次为1号站位(图3b),尾数密度为66.68×103 ind/km2,主要由刀鲚和葛氏长臂虾构成。
2.3 优势种密度空间分布 2.3.1 优势种的种类组成丰水期优势种为安氏白虾和棘头梅童鱼,其重量和尾数所占比例之和分别为35.05%和77.67%。枯水期优势种为葛氏长臂虾、焦氏舌鰨、刀鲚、髭缟虾虎鱼、狭颚绒螯蟹和日本鼓虾,其重量和尾数所占比例之和分别为76.6%和90.73%(表2)。

|
表2 丰、枯水期杭州湾东北部水域渔业资源优势种 Tab. 2 Dominant species of fishery resources in Northeast of Hangzhou Bay during drought and flood seasons |
分别选取丰、枯水期优势度最高的物种作为研究对象。丰水期安氏白虾重量密度最高的站位是2号(图4a),重量密度为153.31 kg/km2。此外,6、7和8号站位也具有较高的重量密度,分别为26.76、39.57和21.02 kg/km2。尾数密度最高的站位也是2号(图4b),尾数密度为255.47×103 ind/km2。其次为7号站位,尾数密度为116.88×103 ind/km2。

|
图4 安氏白虾的密度空间分布 Fig.4 density distribution of Exopalaemon annandalei |
枯水期葛氏长臂虾重量密度较高的站位是1号和10号(图5a),重量密度分别为24.17和24.69 kg/km2。尾数密度最高的站位是10号(图5b),为68.07×103 ind/km2,其次是1号,为35.01×103 ind/km2。此外6号和9号站位也具有较高的尾数密度,分别为13.18×103 ind/km2和14.55×103 ind/km2。

|
图5 葛氏长臂虾的密度空间分布 Fig.5 Density distribution of Palaemon gravieri |
丰水期重量多样性指数(H′)范围在0.36~2.22之间,平均值为1.53±0.61,尾数H′范围在0.16~2.35之间,平均值为1.52±0.62。7、8号站位重量H′最高,分别为2.14和2.22,2、3号站位重量H′最低,分别为0.36和0.92。尾数H′最高是10号站位(2.35),最低是2号站位(0.16),见图6。丰水期尾数H′与尾数密度呈显著相关性(P=0.004<0.05)。

|
图6
丰水期渔业资源多样性指数(H′)的空间分布
Fig.6
Spatial distribution of diversity index (H′) of fishery resources during flood season
a为重量H′,b为尾数H′,图7同。 a means weight H′,b means individual H′,the same in Fig.7. |
枯水期重量H′范围在0.81~2.10之间,平均值为1.66±0.35,尾数H′范围在1.24~2.03之间,平均值为1.61±0.23。10号站位(2.10)重量H′最高,2号站位(0.81)最低。尾数H′最高是7号站位(2.03),10号站位(1.24)最低。此外,1、2和3号站位也具有较低的尾数H′,分别为1.39、1.44和1.46。枯水期尾数H′与尾数密度也呈显著相关性(P=0.024<0.05)。

|
图7 枯水期渔业资源多样性指数(H′)的空间分布 Fig.7 Spatial distribution of diversity index (H′) of fishery resources during drought season |
比较丰、枯水期杭州湾东北部渔业资源密度,丰水期渔业生物重量密度(294.82 kg/km2)和尾数密度(83.19×103 ind/km2)分别大于枯水期重量密度(85.76 kg/km2)和尾数密度(33.88×103 ind/km2)。这一现象的产生与两时期的种类生态习性的差异及主要环境因子变化有一定关系。
丰水期捕获的种类主要由海洋洄游性且个体较大的种类组成,如鮸鱼、白姑鱼和日本蟳等,而枯水期捕获的种类主要由河口定居性且个体较小的种类组成,如焦氏舌鳎、髭缟虾虎鱼和狭颚绒螯蟹等(表1)。相关研究表明[20, 21, 22, 23],杭州湾及其邻近水域是许多海洋洄游性鱼类的索饵场和育肥场。丰水期海洋洄游性鱼类如海鳗、龙头鱼等到近海或河口作产卵洄游和索饵洄游,这势必增加杭州湾东北部渔业生物种类及数量,而枯水期海洋洄游性鱼类多数返回深海越冬,仅剩一些河口定居性种类,如髭缟虾虎鱼、狭颚绒螯蟹等,从而降低了该片海域渔业生物的种类及数量。因此,丰水期的渔业资源密度要大于枯水期。此外,丰、枯水期的生物数量差异也可能与环境因子的变化有关,特别是温度和盐度的改变。温度能影响鱼类的生理活动,较高的水温会促进鱼类的新陈代谢能力与需氧量,从而加快鱼类生长与繁殖[24, 25],较高温度下鱼类的活动范围与觅食水平必然增大。另外,盐度对河口水域鱼类种类丰度和数量分布也有重要影响[26]。从本文研究时期看,丰水期平均温度为(28.3±0.8) ℃,平均盐度为13.4±3.0,而枯水期平均温度仅为(8.5±0.5) ℃,平均盐度为18.1±1.9,两者分别具有显著性差异(P<0.001)。因此,丰水期较高的水温会增加鱼类活动的几率,促使更多鱼类从邻近水域游到杭州湾东北部觅食或产卵,从而增加渔业资源数量及密度。相反,枯水期较高的盐度可能会让一些淡水或河口定居性鱼类“望而却步”,从而减小它们出现在杭州湾东北部的几率。
3.2 优势种数量与群落结构的关系丰水期优势种仅有2种,为安氏白虾和棘头梅童鱼,枯水期优势种多达6种,为葛氏长臂虾、焦氏舌鳎、刀鲚、髭缟虾虎鱼、狭颚绒螯蟹和日本鼓虾,两时期并无共有优势种。这说明杭州湾海域渔业资源群落演替比较剧烈,不同时期渔业资源群落特征明显。从优势种种类及IRI值看,丰、枯水期优势度最高的物种分别是安氏白虾和葛氏长臂虾,均为小型虾类,安氏白虾数量占比达到64.37%,葛氏长臂虾数量占比也接近50%。有研究表明,当优势种由一种或几种个体小、数量大的物种组成时,群落受扰动程度较高,当优势种是由多种数量分布均匀的物种组成时,群落未受扰动或者扰动程度不高[27]。本研究丰、枯水期优势度最高的优势种均是由个体小、数量大的种类组成,说明丰、枯水期杭州湾东北部渔业资源群落受扰动程度均较高,但枯水期拥有更多优势种,说明枯水期渔业生物数量分布较为均匀,群落受扰动程度较丰水期低。
3.3 影响多样性分布的因素多样性指数(H′)常被用来评价水体受到人为影响程度[28],当0 < H′<1,为受到重度影响; H′=1~3,为受到中度影响。本研究范围是0~3,可认为杭州湾东北部海域受到中等程度以上影响。从多样性空间分布看,近岸水域多样性较低,而靠近洋山岛水域多样性较高,这是因为洋山岛属于嵊泗列岛一部分,而岛屿之间底质多样化和水流环境多样化迎合了不同种类海洋生物的生活需求,使得大部分种类生物都有合适的栖息地,特别是一些喜好攀附和贴靠的虾蟹类。反观近岸水域,一是被近岸工业废水排放所污染,二是近岸生态环境多被大堤或者码头所破坏,已经不适合多数海洋生物居住,造成了多样性分布的差异。多样性表征群落的复杂程度,物种多样性的高低。多样性较低说明群落结构不稳定,物种种类组成较为单一。从物种组成看,丰水期近岸多样性较低的站位是2号站位和3号站位,这两个站位均是单一物种占大多数,如2号站位安氏白虾重量占比和尾数占比分别达到91.4%和97.1%,为该站位绝对优势种,3号站位的棘头梅童鱼重量占比也达到78.2%。枯水期多样性较低的站位中(2号站位),刀鲚的重量占比也达到80.6%。这是由于某种物种数量在同一站位中分配不均匀导致多样性指数H′值降低。由此可见,单一优势种的集中出现是导致杭州湾东北部近岸水域多样性指数较低的主要原因,种类数较少仅是次要原因。
| [1] | 罗民波, 陆健健, 沈新强, 等. 大型海洋工程对洋山岛周围海域大型底栖动物生态分布的影响[J]. 农业环境科学学报, 2007, 26(1): 97-102. LUO M B, LU J J, SHEN X Q, et al. Impact of huge ocean engineering on distribution characteristics of macrobenthos in waters around Yangshan islands in Hangzhou bay[J]. Journal of Agro-Environment Science, 2007, 26(1): 97-102. |
| [2] | 章守宇, 邵君波, 戴小杰. 杭州湾富营养化及浮游植物多样性问题的探讨[J]. 水产学报, 2001, 25(6): 512-517. ZHANG S Y, SHAO J B, DAI X J. Studies on eutrophication and phytoplankton diversity in Hangzhou Bay[J]. Journal of Fisheries of China, 2001, 25(6): 512-517. |
| [3] | 蔡燕红. 杭州湾浮游植物生物多样性研究[D]. 青岛: 中国海洋大学, 2006. CAI Y H. The diversity of marine phytoplankton in Hangzhou Bay[D]. Qingdao: Ocean University of China, 2006. |
| [4] | 王晓波, 魏永杰, 秦铭俐, 等. 杭州湾生态监控区浮游动物多样性变化趋势研究[J]. 海洋环境科学, 2008, 27(s1): 67-71. WANG X B, WEI Y J, QIN M L, et al. Studies on variation trend of diversity of zooplankton in ecology monitoring area of Hangzhou Bay[J]. Marine Environmental Science, 2008, 27(s1): 67-71. |
| [5] | 张冬融, 徐佳奕, 徐兆礼, 等. 杭州湾南岸海域春秋季浮游动物分布特征与主要环境因子的关系[J]. 生态学杂志, 2014, 33(8): 2115-2123. ZHANG D R, XU J Y, XU Z L, et al. Distribution pattern of zooplankton in the south part of Hangzhou Bay during spring and autumn in relation to main environmental factors[J]. Chinese Journal of Ecology, 2014, 33(8): 2115-2123. |
| [6] | 周永东, 金海卫, 蒋日进, 等. 浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布[J]. 水产学报, 2011, 35(6): 880-889. ZHOU Y D, JIN H W, JIANG R J, et al. The category composition and abundance distributions of ichthyoplankton along the north-central coast of Zhejiang Province in spring and summer[J]. Journal of Fisheries of China, 2011, 35(6): 880-889. |
| [7] | 陈渊戈. 长江口南支和杭州湾北岸碎波带仔稚鱼群聚特征的比较研究[D]. 上海: 上海海洋大学, 2011. CHEN Y G. Characteristic comparison of fish larval and juvenile assemblages between the surf zones of south branch of Yangtze River estuary and north coast of Hangzhou Bay[D]. Shanghai: Shanghai Ocean University, 2011. |
| [8] | 闫欣, 钟俊生, 王明星, 等. 杭州湾北部水域仔稚鱼种类组成和多样性研究[J]. 上海海洋大学学报, 2014, 23(5): 765-773. YAN X, ZHONG J S, WANG M X, et al. Study on the species composition and diversity of fish larvae and juveniles in the north of Hangzhou Bay[J]. Journal of Shanghai Ocean University, 2014, 23(5): 765-773. |
| [9] | 谢旭, 俞存根, 周青松, 等. 杭州湾海域春、秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(3): 656-663. XIE X, YU C G, ZHOU Q S, et al. Species composition and quantitative distribution of fishes in the Hangzhou Bay during Spring and Autumn[J]. Oceanologia et Limnologia Sinica, 2013, 44(3): 656-663. |
| [10] | 张海波. 杭州湾海洋生物多样性和生态系统健康评价研究[D]. 杭州: 浙江工业大学, 2008. ZHANG H B. Studies on the marine biodiversity and ecosystem health assessment of Hangzhou Bay[D]. Hangzhou: Zhejiang University of Technology, 2008. |
| [11] | 王淼, 张丹, 张玉平, 等. 杭州湾北部张网渔业资源种类组成及群落多样性初步分析[J]. 水产科技情报, 2013, 40(4): 188-192, 198. WANG M, ZHANG D, ZHANG Y P, et al. Preliminary study on Species composition and community diversity of fishery resource by set-net in north Hangzhou Bay[J]. Fisheries Science & Technology Information, 2013, 40(4): 188-192, 198. |
| [12] | 黄海水产研究所. 海洋水产资源调查手册[M]. 2版. 上海: 上海科学技术出版社, 1981. Yellow Sea Fisheries Research Institute. Investigation Handbook of Marine Fisheries Resource[M]. 2nd ed. Shanghai: Shanghai Science and Technology Press, 1981. |
| [13] | SIMPSON E H. Measurement of diversity[J]. Nature, 1949, 163: 688. |
| [14] | 史赟荣, 晁敏, 全为民, 等. 2010年春季长江口鱼类群落空间分布特征[J]. 中国水产科学, 2011, 18(5): 1141-1151. SHI Y R, CHAO M, QUAN W M, et al. Spatial variation in fish community of Yangtze River estuary in spring[J]. Journal of Fishery Sciences of China, 2011, 18(5): 1141-1151. |
| [15] | PINKAS L, OLIPHAMT M S, IVERSON I L K. Food habits of albacore, Bluefin tuna, and bonito in California waters[J]. California Department of Fish and Game, Fish Bulletin, 1971, 152: 1-105. |
| [16] | 吕振波, 李凡, 王波, 等. 黄海山东海域春、秋季鱼类群落结构[J]. 水产学报, 2011, 35(5): 692-699. LV Z B, LI F, WANG B, et al. Community structure of fish resources in spring and autumn in the Yellow Sea off Shandong[J]. Journal of Fisheries of China, 2011, 35(5): 692-699. |
| [17] | 李凡, 张焕君, 吕振波, 等. 莱州湾游泳动物群落种类组成及多样性[J]. 生物多样性, 2013, 21(5): 537-546. LI F, ZHANG H J, LV Z B, et al. Species composition and community diversity of nekton in Laizhou Bay, China[J]. Biodiversity Science, 2013, 21(5): 537-546. |
| [18] | 王蕾, 章守宇, 汪振华, 等. 枸杞岛近岸3种生境鱼类群落组成及岩礁区底栖海藻对鱼类群落结构的影响[J]. 水产学报, 2011, 35(7): 1037-1049. WANG L, ZHANG S Y, WANG Z H, et al. Constitution of fish assemblages in three nearshore habitats and the effect of benthic macroalgae on fish assemblages in Gouqi Island[J]. Journal of Fisheries of China, 2011, 35(7): 1037-1049. |
| [19] | 徐兆礼. 瓯江口海域夏秋季鱼类多样性[J]. 生态学报, 2008, 28(12): 5948-5956. XU Z L. Analysis of fish diversity in the waters off the Oujiang estuary in summer and autumn[J]. Acta Ecologica Sinica, 2008, 28(12): 5948-5956. |
| [20] | 张衡, 朱国平, 陆健健. 长江河口湿地鱼类的种类组成及多样性分析[J]. 生物多样性, 2009, 17(1): 76-81. ZHANG H, ZHU G P, LU J J. Fish species composition and diversity of Yangtze River estuarine wetlands[J]. Biodiversity Science, 2009, 17(1): 76-81. |
| [21] | 张衡, 朱国平. 长江河口潮间带鱼类群落的时空变化[J]. 应用生态学报, 2009, 20(10): 2519-2526. ZHANG H, ZHU G P. Spatiotemporal changes of fish community in Yangtze estuary intertidal zone[J]. Chinese Journal of Applied Ecology, 2009, 20(10): 2519-2526. |
| [22] | 张涛, 庄平, 章龙珍, 等. 长江口中华鲟自然保护区底层鱼类的群落结构特征[J]. 生态学报, 2011, 31(6): 1687-1694. ZHANG T, ZHUANG P, ZHANG L Z, et al. Community structure of demersal fish in Nature Reserve of Acipenser sinensis in Yangtze River estuary[J]. Acta Ecologica Sinica, 2011, 31(6): 1687-1694. |
| [23] | 杨刚, 洪巧巧, 张涛, 等. 长江口中华鲟自然保护区潮间带鱼类群落结构[J]. 生态学杂志, 2012, 31(5): 1194-1201. YANG G, HONG Q Q, ZHANG T, et al. Fish community structure in intertidal area of Acipenser sinensis natural reserve in Yangtze River estuary[J]. Chinese Journal of Ecology, 2012, 31(5): 1194-1201. |
| [24] | 龙华. 温度对鱼类生存的影响[J]. 渔业现代化, 2005(2): 20-22. LONG H. The effect of temperature to fish survival[J]. Fishery Modernization, 2005 (2): 20-22. |
| [25] | 杨阳. 温度和季节对鲤科鱼类游泳、代谢和低氧耐受能力的影响[D]. 重庆: 重庆师范大学, 2014. YANG Y. The effects of temperature and season on the swimming capacity, metabolic and hypoxia tolerance of cyprinid fish[D]. Chongqing: Chongqing Normal University, 2014. |
| [26] | HARRISON T D, WHITFIELD A K. Temperature and salinity as primary determinants influencing the biogeography of fishes in South African estuaries[J]. Estuarine, Coastal and Shelf Science, 2006, 66(1/2): 335-345. |
| [27] | CABRAL H N, COSTA M J, SALGADO J P. Does the Tagus estuary fish community reflect environmental changes?[J]. Climate Research, 2001, 18(1/2): 119-126. |
| [28] | 国家环保局《水生生物监测手册》编委会. 水生生物监测手册[M]. 南京: 东南大学出版社, 1993. National Environmental Protection Agency《Aquatic Monitoring Handbook》Editorial Committee. Aquatic Monitoring Handbook [M]. Nanjing: Northeastern University Press, 1993. |