 2016, Vol. 25
2016, Vol. 25

2. 上海海洋大学 水域环境生态上海高校工程研究中心, 上海 201306;
3. 上海海洋大学 海洋科学学院, 上海 201306
温室气体的控制大概是最具有挑战性的环境政策问题。二氧化碳(CO2)在全球性长期稳定的温室气体中含量最高,占总数的63%以上,也是导致全球变暖的主要温室气体[1, 2]。
海洋对调节全球气候至关重要,特别是对减缓温室效应的影响起着巨大的作用,通过物理溶解、海水碳酸盐系统缓冲作用和海洋浮游植物光合作用吸收大气中的CO2[3],同时,海水中CO2体系影响着海洋中许多化学平衡。目前人类活动每年排放的CO2以碳计为55亿吨,其中海洋吸收了人类排放CO2总量的30%~50%[4],所以区域性以及全球海洋是吸收(汇)还是排放(源)CO2就显得非常重要[5]。
我国的海水养殖已经居世界首位,其中贝藻养殖活动可能成为减排增汇的有效手段[6],有关养殖海域海水中的pCO2的变化特征与CO2交换通量已进行了一些研究报道。张继红等[7]于2011-2012年对桑沟湾表层水CO2体系各参数进行研究,分析了该海域pCO2的季节变化可能与物理、生物和大规模贝藻养殖之间的相关性。蒋增杰等[8]对山东俚岛湾养殖海域海-气界面CO2交换通量进行估算,指出栽培大型海藻促进了海洋对大气中CO2的吸收。王文松等[9]于2011年春季对胶州湾春季表层海水pCO2和海-气界面CO2交换通量进行测量时得出该海域表现为大气中CO2强汇,生物活动是影响海水pCO2的主要因素。
福建省三沙湾总面积为714 km2,水域开阔,仅东南方经东冲口与东海相通,是我国南方典型的近海封闭型海湾,湾内咸淡水交混,营养盐丰富,具有独特的海洋生态环境,不仅是全国唯一的内湾性大黄鱼产卵场与最大的海水网箱养殖基地[10, 11],同时还有大型海藻、贝类和海参等养殖种类,盐田港是三沙湾重要港湾组成之一。然而,对该海湾的CO2的源汇问题的研究尚未进行。本研究根据2012-2013年4个调查航次的调查数据,估算了该养殖水域的海-气界面CO2交换通量,并对影响其时空变化的环境因子进行分析,为我国封闭型海湾海水养殖业的可持续发展提供科学依据。
1 材料与方法 1.1 研究海域概况福建省三沙湾盐田港主要养殖大黄鱼(Pseudosciaena crocea)、长牡蛎(Crassostrea gigas)和海参(Sea Cucumber),根据季节更替栽培龙须菜(Gracilaria lemaneiformis)和海带(Laminaria japonica)。鱼类网箱养殖规模为1.15×104个网箱,鱼类的饵料主要是冰鲜小杂鱼,养殖周期通常是2~3年。除了鱼类,长牡蛎养殖规模是 3.37 km2,养殖海参约1 100 口,总产量约160 t。大型海藻龙须菜和海带是本地在不同季节养殖的主要物种,9月份至来年2月份主要栽培龙须菜,海带大规模栽培时间是12月底至次年5月份,调查期间两者栽培规模分别是72.44 km2和181.89 km2。
1.2 采样时间、站位及方法本研究分别于2012年11月和2013年2月、5月、8月共4个航次在福建省三沙湾内的盐田港(26.72° ~ 26.84°N,119.76°~ 119.83°E)10个站位进行调查(图1)。其中,1号站位位于非养殖区,2号和5-10号位于大型海藻养殖区内,3号位于长牡蛎养殖区内,4号位于大黄鱼网箱养殖区内。

|
图1 盐田港调查站位图 Fig.1 Location of sampling sites in Yantian Harbor |
样品均按《海洋监测规范》规定的方法采集、处理和保存。在每个调查站位应用Niskin采水器采集表层(水面下0.5 m)3个水样作为重复,迅速导入500 mL磨口玻璃瓶中,保存在4 ℃冰箱中备用。现场应用YSI多参数水质分析仪测定pH、表层水温(SST)和盐度(S),应用碘量法测定溶解氧(DO)含量(GB 17378.4—2007)。将500 mL水样经0.45 μm醋酸纤维滤膜过滤后冷冻保存带回实验室,经丙酮萃取后应用Turner荧光仪测定叶绿素a(Chl.a)的含量。总碱度(TA)采用pH法测量(GB12763.4—2007),溶解无机碳(DIC)应用日本岛津总有机碳分析仪(TOC-VCPH)进行测定。
1.3 CO2交换通量和无机碳体系各分量浓度估算方法海-气界面O2交换通量的计算公式[8]为

假定海水中CO2体系处于平衡,如果海水的温度、盐度和压力已经测定,那么pH、总碱度(TA)、溶解无机碳(DIC)和表层水二氧化碳分压(pCO2)就可以任意测定两个,然后计算出其余的参数[15, 16]。本文利用获得pH和TA数据,计算CO2体系的其他参数。
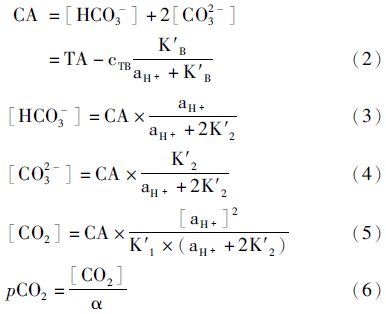
所有数据均用平均值±标准误表示,应用SPSS 13.0软件对不同季节和不同站位的无机碳体系各分量的浓度、pCO2和CO2交换通量进行双因素方差分析(ANOVA),当P<0.01时为差异极显著,当P<0.05时为差异显著,数据的多重比较采用SPSS软件中的LSD法及Duncan氏法进行分析。分布图中的绘制采用Surfer 8.0软件。
2 结果 2.1 海区水文环境特征三沙湾盐田港表层水温四季变化明显,调查海域春季和夏季水温较高,平均值分别为(23.38±0.67) ℃和(29.97±0.08) ℃,水温由湾内上游到下游逐渐降低;秋季水温逐渐下降,变化范围在16.61~19.71 ℃之间,冬季表层水温仅有(12.96±0.27)℃。夏季pH最低,为7.53±0.14,上游站位偏低,其他3个季节是大型海藻栽培的季节,pH有所升高,其中冬季pH达到最高,为7.87±0.05,春季和秋季pH分别7.71±0.17和7.75±0.03,在这3个季节中,位于下游的海藻栽培区pH高于其他功能区。海水表层盐度具有显著的四季变化,表现为上游到下游逐渐增加的分布特征。夏季盐度最高,平均值为26.66±1.60,春季盐度最低,平均值仅有19.03±4.51,其中,位于上游的1号站位盐度全年最低只有10.30,秋季和冬季变化范围分别为20.03~25.72和19.73~24.18。该海域叶绿素a(Chl.a)由春季到冬季逐渐降低,变化范围在1.12~3.92 μg/L之间,夏季5号和6号站位Chl.a高于上游和下游的其他站位,最高达3.61 μg/L。溶解氧(DO)冬季最高,为(8.14±0.12) mg/L,夏季最低,平均值是(6.49±0.07) mg/L,春季和秋季为(7.08±0.28) mg/L和(7.33±0.04) mg/L,呈现海藻养殖区>非养殖区>贝类物养殖区>网箱养殖区的分布特征(表1)。

|
表1 不同季节表层海水基本参数 Tab.1 The basic parameters of surface water in different seasons |
调查期间,三沙湾盐田港表层海水中无机碳体系各分量浓度在不同季节间差异极显著。DIC和HCO3-浓度年变化范围分别为955.00~1 957.08和905.08~1 848.13 μmol/L,平均值为(1 628.74±142.84) μmol/L和(1 536.14±109.58)μmol/L,其中HCO3-是其重要的组成部分,占全年DIC平均值的94.32%,秋、冬季节表层海水的HCO3-浓度比春、夏季节高。海水中CO32-年变化范围为10.14~124.78 μmol/L,平均值为(70.16±16.42)μmol/L,该海域夏季CO32-浓度显著高于其他3个季节;CO2浓度的季节变化在11.48~39.78 μmol/L之间,均值为(22.44±10.51)μmol/L,与CO32-浓度季节变化相反,夏季CO2浓度低于其他3个季节,仅为3个季节浓度均值的58.25%(表2)。同一季节CO2和CO32- 的浓度在不同站位之间差异显著和极显著,而DIC 和HCO3- 浓度在同一季节的不同站位之间的差异不显著(表3)。

|
表2 不同季节表层海水无机碳体系各分量浓度 Tab.2 DIC concentration and its component forms in different seasons μmol/L |

|
表3 不同季节表层海水无机碳体系各分量的双因素方差分析 Tab.3 Two-way ANOVA tests of CO2 system in different seasons |
盐田港表层海水中pCO2的平面分布如图2所示,由于养殖海域水环境系统的复杂性,该海域海水中pCO2的分布不均匀,四季变化显著。调查期间,表层海水中pCO2的年变化范围为391.27~1 200.49 μatm,平均值为(6 52.71± 51.73)μatm。春季和秋季pCO2的平均值分别为(744.44±62.56) μatm和(724.66±58.41)μatm,差异不显著(P=0.740);冬季与夏季之间差异不显著,平均值为(572.99±32.63)μatm和(561.89± 28.37)μatm,显著低于秋季和春季(P冬秋=0.015、P冬春=0.006、P夏秋=0.009和P夏春=0.004),见表4。海水pCO2的值在同一季节的不同站位之间的差异显著(P=0.039,表5),在不同季节之间的差异极显著(P=0.001)。秋、冬、春3个季节大型海藻生长旺盛,下游大规模栽培的海藻活动加剧,吸收海水中大量无机碳,使得pCO2偏低;从图2可知这3个季节的表层海水中pCO2分布趋势相同,表现为上游向下游不断降低;夏季是海藻栽培的空白期,5号和6号站位较高的Chl.a含量佐证了中游区域浮游植物活动加剧,pCO2分布表现为中部区域偏低,向上游和下游呈辐射状增加的趋势。

|
图2 盐田港表层海水pCO2分布(μatm) Fig.2 Distribution of sea surface pCO2 in Yantian Harbor (μatm) |

|
表4 不同季节表层海水pCO2 Tab.4 Sea surface pCO2 in different seasons μatm |

|
表5 不同季节海水pCO2的方差分析 Tab.5 Two-way ANOVA tests of sea surface pCO2 in different seasons |
盐田港的海-气界面CO2交换通量的季节变化显著,全年海-气界面CO2交换通量范围在 0.25~6.93 μmol/(m2·d)之间,平均值为(2.54±0.92)μmol/(m2·d),表现为大气CO2的源。盐田港秋、春季海-气界面CO2交换通量的均值为(3.37±0.88) μmol/(m2·d)和(3.12±1.85)μmol/(m2·d),差异不显著(P=0.553),冬季平均值为(2.304±0.72)μmol(/m2·d),显著低于秋季(P=0.029),但与春季差异不显著(P=0.102);夏季显著低于秋、春和冬季(P=0.000、P=0.001和P=0.043)见表6、7。根据图3可知,在秋、冬、春3个季节中,盐田港CO2交换通量均表现为上游向下游不断降低,下游海藻栽培区表现的源弱于3号长牡蛎养殖区和4号网箱养殖区;在夏季表现为从中部区域向上游和下游辐射状增加的趋势,除了10号站位,位于上游的长牡蛎养殖区和网箱养殖区表现的源要强于下游的非养殖区。

|
表6 不同季节海-气界面CO2交换通量F Tab.6 Sea-air CO2 flux in different seasons |

|
表7 不同季节海-气界面CO2交换通量的方差分析 Tab.7 Two-way ANOVA tests of sea-air CO2 flux in different seasons |

|
图3 盐田港海-气界面CO2交换通量分布(μatm) Fig.3 Distribution of sea-air CO2 flux (μatm) in Yantian Harbor |
由表8可知,从全年的尺度来看,海-气界面CO2交换通量与pH极显著负相关,与盐度(S)和总碱度(TA)显著负相关,而与其他环境因子不相关。从不同季节来看,CO2交换通量与4个季节的pH显著和极显著负相关;表层水温(SST)和盐度(S)影响冬、春季节CO2交换通量的环境因子;DIC和总碱度(TA)是影响春、夏季节CO2交换通量的环境因子;另外,春季CO2交换通量与DO含量显著负相关,而各季节的Chl.a浓度与CO2交换通量均无显著的相关性。

|
表8 CO2交换通量与水环境因子的相关系数 Tab.8 Pearson correlation coefficients between sea-air CO2 flux and environmental factors |
三沙湾为我国南方位于福建省东北部沿海的一个复杂的海湾[17],盐田港位于三沙湾内湾之中。本研究结果表明,盐田港表层海水DIC浓度低于我国北方典型海湾桑沟湾和乳山湾的DIC浓度[4, 8],这可能与盐田港所在的地理区位与水文条件有关。研究表明,表层水pCO2受到各种物理、化学、生物条件的影响,如水温、盐度、深度、pH、N、P、Chl.a、PP等[7]。其中,物理过程如水温、盐度是重要的影响因素之一[18]。盐田港受到陆源输入的影响,盐度由上游到下游逐渐增加,海水中离子浓度加大,导致海水中碳酸盐体系电离度下降,CO2在海水中的溶解度随之降低,与表层海水pCO2由上游到下游降低的分布趋势相一致。
宋金明[5]报道温带海域表层水pCO2和水温之间存在着显著的正相关性。但近岸养殖生态系统的复杂性使得pCO2的分布存在着较大的不均匀性[7],不同于大洋上pCO2的分布[19]。生物条件可能是影响三沙湾表层水pCO2空间分布的主要生态因子[7, 8]。夏季,表层海水温度达到全年最高,CO2的溶解度随之下降,有利于CO2从水体进入大气中;盐田港夏季并无大型海藻类栽培,但随着水温和营养盐浓度的升高,浮游植物的现存量(2.72×104~16.23×104 cells/L)显著高于其他季节,浮游植物光合作用会降低海水中的pCO2[20],这可能是8月份盐田港海水表层部分监测站位的pCO2低于其他季节的原因。
盐田港每年从10月份开始大规模进行龙须菜养殖,至12月份开始陆续更换为海带,栽培时间持续到来年的5月份。盐田港龙须菜和海带栽培的区域主要集中在盐田港的湾口区域,龙须菜和海带等大型海藻通过光合作用将海水中的溶解无机碳转化为有机碳,吸收水体中的DIC,使得水体中pCO2有所降低。因此,秋季(11月)、冬季(2月)和春季(5月)表层海水pCO2的空间分布表现为盐田港湾口海藻养殖区低于湾内长牡蛎和网箱养殖区域。盐田港夏季(8月)无大规模海藻栽培,下游表层海水pCO2的平均值与部分站位的pCO2值低于秋、冬和春季的相应区域。贝类生长过程中通过呼吸作用和钙化作用释放CO2的量高于本海域的初级生产力,因此四季中长牡蛎养殖区pCO2处于较高水平[21]。4号网箱养殖区在春季和夏季pCO2低于邻近的贝类养殖区和非养殖区,鱼类排泄和残饵使得该区域水体营养盐丰富,此期间该功能区Chl.a在3.42~8.62 μg/L之间,浮游植物的光合作用为降低pCO2做出主要贡献。
3.2 海-气界面O2交换通量的季节变化与分析盐田港四个季节均表现为大气CO2的弱源,而北方的桑沟湾、乳山湾、大连湾以及胶州湾的大部分区域均表现为大气CO2的汇[4, 8, 22, 23]。虽然大型海藻和浮游植物会通过光合作用降低海水pCO2,但这表明有其他无机碳源的输入。一方面,大规模养殖的长牡蛎在形成贝壳的钙化过程中与呼吸作用协同释放CO2[4],同时,过于密集的网箱养殖产生残饵和粪便,鱼类生长代谢过程中产生CO2[11];另一方面两岸村镇农业与生活污染物输入以及沉积物溶解释放,加之大型海藻的栽培量较少,使得三沙湾盐田港养殖海域表现为CO2的弱源。盐田港夏季海-气界面O2交换通量显著低于其他季节,这可能与夏季DIC输入减少和浮游植物较强烈的光合作用有关[24]。另外,湾口区域与湾内区域的水文动力条件的差异也是影响盐田港海-气界面O2交换通量时空差异的原因之一,但还有待于进一步研究。
温带海域表层海水pCO2与水温之间存在着显著的正相关关系[15]。盐田港4个季节的航次调查,只有冬、春季pCO2与这种普遍的规律相一致。盐田港春季海-气界面O2交换通量与DIC和水体DO的含量显著相关,表明水体的物理过程和生物活动的耦合作用造成pCO2的变化。通常,初级生产者的光合作用是影响DO含量的主要原因。5月份,盐田港大规模的海带和龙须菜栽培同时存在,同时随着水温升高和营养盐浓度的增加,浮游植物的生物量也增加(Chl.a的最高值为9.62 μg/L),因此春季初级生产者的光合作用可能是影响盐田港海-气界面O2交换通量的主要原因之一。盐田港夏季海-气界面O2交换通量与pH、DIC和TA显著相关外,与DO并无显著的相关关系。虽然浮游植物光合作用是影响盐田港夏季海水表层的pCO2的主要因素之一,但其他来源的DIC输入可能也是影响海-气界面O2交换通量的重要生态要素。总体来看,与环境因子之间关系的复杂性表明CO2交换通量是受到养殖生物、浮游生物的生理生态过程与海域物理过程耦合作用的结果。
本研究结果对评价海水养殖海域生态环境状况和指导海水养殖可持续发展都具有一定的指导意义。调查期间,海藻大规模栽培的季节CO2交换通量与长牡蛎养殖区、网箱养殖区和夏季无海藻栽培区相比,表现的大气CO2源偏弱。龙须菜和海带的轮换栽培吸收水体中的碳,减弱表层海水的pCO2,提高养殖海域生态系统吸收CO2的能力。GAO等[25]研究认为大型海藻不仅吸收水体中的CO2,同时对HCO3-具有较强的吸收能力,能直接吸收HCO3-作为外在光合作用的碳源。可见开展多营养层次的综合养殖可在一定程度上调节海产经济动物养殖区海-气界面O2交换通量。本文碳酸盐参数的计算前提是基于碳酸盐体系的化学平衡,而在浅海养殖区域水体中碳酸盐体系有时会处于非平衡状态[7]。因此,应用化学平衡方法估算海湾养殖区pCO2和CO2通量有一定的“不确定性”和局限性,这将在今后的研究中重点解决。
| [1] | LI L, ZHAO N, WEI W, et al. A review of research progress on CO2 capture, storage, and utilization in Chinese Academy of Sciences [J]. Fuel, 2013, 108: 112-130. |
| [2] | DE_RICHTER R K, MING T Z, CAILLOL S. Fighting global warming by photocatalytic reduction of CO2 using giant photocatalytic reactors [J]. Renewable and Sustainable Energy Reviews, 2013, 19: 82-106. |
| [3] | 宋金明. 二氧化碳的温室效应与全球气候及海平面的变化[J]. 自然杂志, 1991, 14(9): 649-653. SONG J M. The CO2 greenhouse effect and global climate and sea level changes [J]. The Journal Nature, 1991, 14(9): 649-653. |
| [4] | 蒋增杰, 方建光, 王巍, 等. 乳山宫家岛以东牡蛎养殖水域秋季海-气界面CO2交换通量研究[J]. 水产学报, 2012, 36(10): 1592-1598. JIANG Z J, FANG J G, WANG W, et al. Sea-air CO2 flux in Crassostrea gigas aquaculture area of East Gongjia Island, Rushan, Shandong, in autumn [J]. Journal of Fisheries of China, 2012, 36(10): 1592-1598. |
| [5] | 宋金明. 海洋碳的源与汇[J]. 海洋环境科学, 2003, 22(2): 75-80. SONG J M. Carbon sources and sinks in oceans [J]. Marine Environmental Science, 2003, 22(2): 75-80. |
| [6] | TANG Q S, ZHANG J H, FANG J G. Shellfish and seaweed mariculture increase atmospheric CO2 absorption by coastal ecosystems [J]. Marine Ecology progress Series, 2011, 424: 97-105. |
| [7] | 张继红, 吴文广, 任黎华, 等. 桑沟湾表层水pCO2的季节变化及影响因素分析[J]. 渔业科学进展, 2013, 34 (1): 57-64. ZHANG J H, WU W G, REN L H, et al. Seasonal variation of pCO2 and its potential influencing factors in aquaculture areas of Sanggou Bay [J]. Progress in Fishery Sciences, 2013, 34(1): 57-64. |
| [8] | 蒋增杰, 方建光, 韩婷婷, 等. 大型藻类规模化养殖水域海-气界面CO2交换通量估算[J]. 渔业科学进展, 2013, 34(1): 50-56. JIANG Z J, FANG J G, HAN T T, et al. Estimation of sea-air CO2 flux in seaweed aquaculture area, Lidao Bay [J]. Progress in Fishery Sciences, 2013, 34(1): 50-56. |
| [9] | 王文松, 薛明, 张龙军. 2011年3月胶州湾表层海水pCO2及海-气界面通量[J]. 中国海洋大学学报:自然科学版, 2012, 42(7/8): 144-149. WANG W S, XUE M, ZHANG L J. Distribution and controlling factors of sea surface partial pressure of CO2 and Air-Sea CO2 flux in Jiaozhou Bay during March, 2011[J]. Periodical of Ocean University of China, 2012, 42(7/8): 144-149. |
| [10] | 蔡清海. 福建三沙湾海洋生态环境研究[J]. 中国环境监测, 2007, 23(6): 101-105. CAI Q H. Study on Maine ecological environment of Sansha Bay in Fujian[J]. Environmental Monitoring in China, 2007, 23(6): 101-105. |
| [11] | 马祖友, 夏永健, 石志洲, 等. 2011年三沙湾增养殖区水环境质量评价[J]. 海洋开发与管理, 2013(7): 75-78. MA Z Y, XIA Y J, SHI Z Z, et al. The assessment of water environmental quality in the mariculture area of Sansha Bay in 2011 [J]. Marine Development and Management, 2013(7): 75-78. |
| [12] | TSUNOGAI S, WATANABE S, SATO T. Is there a “continental shelf pump” for the absorption of atmospheric CO2 [J]. Tellus B, 1999, 51(3): 701-712. |
| [13] | 宋金明. 中国近海生物地球化学[M]. 济南: 山东科学技术出版社, 2004. SONG J M. China Offshore Biogeochemistry[M]. Jinan: Shandong Science and Technology Press, 2004. |
| [14] | 陈佳荣. 水化学[M]. 北京: 中国农业出版社, 1996. CHEN J R. Water Chemistry[M]. Beijing: China Agriculture Press, 1996. |
| [15] | MILLERO F J. Thermodynamics of the carbon dioxide system in the oceans[J]. Geochimica et Cosmochimica Acta, 1995, 59(4): 661-677 |
| [16] | MEHRHACH C, CULLBERSON C H, HAWLEY J E, et al. Measurement of the apparent dissociation constants of carbonic acid in seawater of at atmospheric pressure[J]. Limnology and Oceanography, 1973, 18(6): 897-907 |
| [17] | 中国海湾志编纂委员会, 中国海湾志[M]. 北京: 海洋出版社, 1994. The gulf records compilation committee of China, The gulf records[M]. Beijing: Marine Press, 1994. |
| [18] | WEISS R F. Carbon dioxide in water and seawater: the solubility of a non-ideal gas [J]. Marine chemistry, 1974, 2(3): 203-215. |
| [19] | COOPER D J, WATSON A J, LING R D. Variation of pCO2 along a North Atlantic shipping route (U. K. to the Caribbean): A year of automated observations [J]. Marine Chemistry, 1998, 60(1/2): 147-164. |
| [20] | 张龙军, 王婧婧, 张云, 等. 冬季北黄海表层海水pCO2分布及其影响因素探讨[J]. 中国海洋大学学报, 2008, 38(6): 955-960. ZHANG L J, WANG J J, ZHANG Y, et al. Distribution and controlling factors of surface seawater partial pressure of CO2 in the Northern Yellow Sea during winter [J]. Periodical of Ocean University of China, 2008, 38(6): 955-960. |
| [21] | MARTIN S, THOUZEAU G, CHAUYAU D, et al. Respiration, calcification, and excretion of the invasive slipper limpet, Crepidula fornicate L.: Implications for carbon, carbonate, and nitrogen fluxes in affected areas[J]. Limnology and Oceanography, 2003, 51(5): 1996-2007. |
| [22] | 刘启珍, 张龙军, 薛明. 胶州湾秋季表层海水pCO2分布及水-气界面通量[J]. 中国海洋大学学报, 2010, 40(10): 127-132. LIU Q Z, ZHANG L J, XUE M. Distribution and controlling factors of surface seawater partial pressure of CO2 and Air-Sea carbon fluxes in Jiaozhou Bay during autumn [J]. Periodical of Ocean University of China, 2010, 40(10): 127-132. |
| [23] | 嵇晓燕, 崔广柏, 杨龙元, 等. 太湖水-气界面CO2交换通量观测研究[J]. 环境科学, 2006, 27(8): 1479-1486. JI X Y, CUI G B, YANG L Y, et al. Measurement of the CO2 flux on the water-air interface of Taihu Lake [J]. Environmental Science, 2006, 27(8): 1479-1486. |
| [24] | 曲宝晓, 宋金明, 袁华茂, 等. 东海海-气界面二氧化碳通量的季节变化与控制因素研究进展[J]. 地球科学进展, 2013, 28(7): 783-793. Qu B X, SONG J M, YUAN H M, et al. Advances of seasonal variations and controlling factors of the sea-air CO2 flux in the East China Sea [J]. Advances in Earth Science, 2013, 28(7): 783-793. |
| [25] | GAO K S, MCKINLEY K R. Use of macroalgae for marine biomass production and CO2 remediation: a review[J]. Journal of Applied Phycology, 1994, 6(1): 45-60. |
 , HAN Hongbin1, HU Ming1, WU Hailong1, ZHANG Jianheng3, HUO Yuanzi1, 2
, HAN Hongbin1, HU Ming1, WU Hailong1, ZHANG Jianheng3, HUO Yuanzi1, 2 , HE Peimin1, 2
, HE Peimin1, 2 2. Water Environment and Ecology Engineering Center of Shanghai Institute of Higher Education, Shanghai Ocean University, Shanghai 201306, China;
3. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China