 2016, Vol. 25
2016, Vol. 25



虾类在大洋和近岸食物网中皆为重要的饵料生物,其本身亦是人类蛋白质来源的主要海洋生物类群之一,对其组成和分布的研究有助于我们更全面地认识海洋生态系统中的食物网结构,并促进虾类资源的利用和保护工作。
国外学者围绕虾类的研究主要从生物学和群落生态学两个方面进行探讨,主要包括种类组成、优势种、多样性和群落结构等内容,GONZALEZ-GORDILLO等[1]比较了3个海岸带甲壳类幼体群体的季节和空间分布特征,STEPHANIE等[2]分析了人工防御设施对十足目甲壳类的影响,MARCO和ANGELO[3]对撒丁岛海峡中部半深海区域底栖甲壳类的群落结构特征进行了研究,COSTA等[4]探讨了环境因子对巴西东南部海域阿根廷虾(Artemesia longinaris)丰度以及时空分布的影响。迄今为止,我国在虾类群落生态学研究方面已经积累了大量资料。国内学者十分重视虾类生态学、区系和空间分布等方面的研究,如宋海棠等和陈小庆等[5, 6, 7]学者研究了东海大陆架海域经济虾类组成、分布规律及群落结构区系特征等。近岸、河口等中小尺度的虾类群落研究亦较为丰富,如齐海明等[8]对椒江口海域春秋季虾类的群落结构特征进行了分析;徐兆礼等[9]对瓯江口海域虾类资源分布特征进行了研究。
稳定且多样的生物群落往往是近岸各种典型栖息地结构完整和功能健康的标志,尤其在岛礁海域,复杂的栖息地结构和多样化的生源要素成为虾类生存和繁衍的良好生境。纵观已有研究,一些已经成为海洋保护区或者特别保护区的岛礁区域,其虾类群落生态学也逐渐得到关注,已有的报道有谢汉阳等[10]对中街山水域虾类组成及其群落多样性进行了分析,晁文春[11]等探讨了春夏季南麂列岛海域甲壳类种类组成及分布特征等。针对马鞍列岛海洋特别保护区以及内部岛屿如东南部枸杞岛海域潮下带、岩礁区域、海藻场及其近岸海域鱼类、底栖生物和浮游植物等方面的研究已有很多[12, 13, 14],至今未见马鞍列岛海域虾类群落方面的专门报道。作为浙江省重要的海洋牧场建设示范区,查清海域底栖虾类的群落现状非常迫切,深入研究岛礁生态系统各基础环节的宏观状况是保护区开发和管理的直接依据。本文的目的就是要探明马鞍列岛海洋保护区虾类群落组成特征,以期丰富我国岛礁水域虾类群落生态学研究内容,为保护区的生物资源管理和保护以及生态修复等提供参考。
1 材料与方法 1.1 研究区域与站点布设马鞍列岛海洋特别保护区位于浙江舟山群岛北部(122°35′-l22°51′E;30°41′-30°52′N),是东海海洋保护区中面积最大的(549 km2,图1)。沿岸冲淡水、黄海冷水团、黑潮暖流的分支台湾暖流表层水和台湾暖流深层水在此交汇[15],加上研究海域大大小小散落着众多岛屿,形成水文要素如温度、盐度等时空变化极为复杂的海域环境,同时诸多水系的存在也使得研究海域饵料生物组成较为丰富,为许多经济鱼虾蟹类索饵育肥、繁殖提供了良好的生态栖息环境。

|
图1 马鞍列岛海域虾类拖网采样站位分布 Fig.1 Distribution of shrimps sampling stations in Ma′an Archipelago area “●”采样站位;“×”定置网具分布区域。 “●”sampling stations; “×”the area with setting nets. |
对马鞍列岛海域进行网格化,以1′×1′的范围为采样网格。在不受捕捞作业、海上工程和海水养殖影响的网格内进行随机采样。
1.2 采样方法和样品处理2013年夏季(7月)、秋季(10月)和2014年冬季(3月)、春季(5月)共在马鞍列岛海域进行了4个航次调查。租用40 hp的桁杆虾拖网船(浙渔67127)作为采样船,该船长15 m,宽2.7 m,吃水0.5 m。调查网具为桁杆虾拖网,桁杆长15.5 m,直径10 cm;网口扩张高度1 m,网袖网目32 mm,网囊网目为20 mm。为降低对小区域内的捕捞干扰,每一采样站点拖曳时间控制在20 min,平均拖速1.0 kt。
渔获总重大于20 kg时,随机抓取其中的样品不少于10 kg,并将少见种和偶见种全部取出;渔获总重小于20 kg时,则全部取样。种类鉴定到其分类的最低阶元。并对每一个体进行称重、计数及常规生物学指标(如头胸甲长,体长,性别和摄食等级等)的测量。各项测定皆依照《海洋生物生态调查技术规程》进行。各种类生态类群的确定依据宋海棠等著《东海经济虾蟹类》。在虾类拖网调查的同时,采用CTD测定各站位的水深、温盐等环境因子。
1.3 数据处理和统计分析依据扫海面积法[16]将虾类丰度和生物量数据统一换算为单位面积的渔获量,公式如下:

运用相对重要性指数(IRI)[18]、Margalef种类丰富度指数(D)[19]、Shannon-Wiener多样性指数(H′)[20]和Pielou均匀度指数(J′)[21]对虾类优势种组成和多样性进行分析,公式依次如下:
相对重要性指数(IRI):
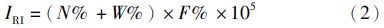
Margalef种类丰富度指数(D):

Shannon-Wiener多样性指数(H′):

Pielou均匀度指数(J′):

参考方差分析贡献方法分析虾类密度变化的动力学,以虾类总生物量或总丰度为因变量,各优势种生物量或丰度为自变量,分析优势种对虾类数量变化的影响 [22]。采用SPSS 19.0对各季节的多样性指数进行单因素方差分析,显著水平均为α=0.05。
2 结果与分析 2.1 马鞍列岛海域虾类种类组成2013年7月至2014年5月在马鞍列岛海域共采集虾类20种,隶属于2目9科16属(表1)。其中对虾科的种类数最多(7种),占总种类数的35%;其次为藻虾科(4种);长臂虾科和鼓虾科各2种;虾蛄科、樱虾科、长额虾科、玻璃虾科和褐虾科各1种。虾类种类数夏季最少,仅12种,隶属于6科11属;秋季最多,为16种,隶属于8科13属;冬季采获13种,隶属于8科10属,春季采获14种,隶属于8科12属。

|
表1 马鞍列岛海域虾类种类组成 Tab. 1 Species composition of the shrimps in Ma′an Archipelago area |
马鞍列岛海域虾类种类组成中,葛氏长臂虾(Palaemon gravieri)、细巧仿对虾(Parapenaeopsis tenella)、哈氏仿对虾(Parapenaeopsis hardwickii)、口虾蛄(Oratosquilla oratoria)、日本鼓虾(Alpheus japonicus)、鲜明鼓虾(Alpheus distinguendus)、细螯虾(Leptochela gracilis)、鞭腕虾(Lysmata vittata)和水母虾(Latreutes mucronatus)在四个季节均有出现;中华管鞭虾(Solenocera crassicornis)和中国毛虾(Acetes chinensis)在三个季节出现;鹰爪虾(Trachypenaeus curvirostris)、周氏新对虾(Penaeus joyneri)、脊腹褐虾(Crangon affinis)和戴氏赤虾(Metapenaeopsis dalei)仅出现在某两个季节;其他种类仅出现在单个季节。
2.2 马鞍列岛海域虾类优势种组成和季节变化夏季虾类优势种有5种,依次为葛氏长臂虾、细巧仿对虾、中华管鞭虾、口虾蛄和哈氏仿对虾,占该季节种类数的41.7%。葛氏长臂虾和细巧仿对虾的相对重要性指数明显高于其他种类(表2),生物量合占虾类总生物量的65.8%,丰度合占虾类总丰度的77.4%。其中,细巧仿对虾的生物量优势较高,为72.4 kg/km2,而葛氏长臂虾的丰度优势较高,为60.9×103 ind/km2。细螯虾、日本鼓虾、水母虾和鲜明鼓虾为常见种。

|
表2 马鞍列岛海域虾类主要优势种生态特征 Tab. 2 Ecological character of main dominant species of shrimps in Ma′an Archipelago area |
秋季虾类优势种有7种,依次为葛氏长臂虾、口虾蛄、中华管鞭虾、细巧仿对虾、哈氏仿对虾、鞭腕虾和日本鼓虾,占该季节种类数的43.8%。葛氏长臂虾和口虾蛄的相对重要性值明显高于其他种类,其生物量合占虾类总生物量的59.7%,丰度合占虾类总丰度的48.9%。其中,口虾蛄的生物量优势较高,为20.0 kg/km2,而葛氏长臂虾的丰度优势较高,为11.0×103 ind/km2。中华管鞭虾丰度比例高于口虾蛄,也是较重要的优势种类。水母虾、鲜明鼓虾和细螯虾为常见种。
冬季虾类优势种有9种,依次为葛氏长臂虾、细巧仿对虾、水母虾、细螯虾、口虾蛄、中国毛虾、鲜明鼓虾、脊腹褐虾和日本鼓虾,占该季节种类数的69.2%。葛氏长臂虾的相对重要性值最高,其生物量比例达54.5%,为30.4 kg/km2。细巧仿对虾和水母虾的相对重要性值仅次于葛氏长臂虾,占丰度优势较高,其丰度总和占虾类总丰度的51.1%,分别为16.3×103 ind/km2和14.3×103 ind/km2。鞭腕虾、哈氏仿对虾和安氏白虾为常见种。
春季虾类优势种有5种,依次为细巧仿对虾、水母虾、葛氏长臂虾、口虾蛄和日本鼓虾,占该季节种类数的35.7%。细巧仿对虾的相对重要性指数明显高于其他种类,其生物量和丰度比例均最高,分别为35.7 kg/km2和48.1×103 ind/km2。水母虾的丰度比例略低于细巧仿对虾,为40.1×103 ind/km2,也是重要的优势种类。中国毛虾、哈氏仿对虾、细螯虾和周氏新对虾为常见种。
葛氏长臂虾、细巧仿对虾和口虾蛄为虾类全年优势种,其中葛氏长臂虾在夏、秋和冬季为绝对优势种,细巧仿对虾在春季为绝对优势种;日本鼓虾在秋、冬和春季三个季节为优势种;哈氏仿对虾、中华管鞭虾和水母虾在某两个季节成为优势种;鞭腕虾、细螯虾、中国毛虾和脊腹褐虾仅为某单个季节的优势种。
2.3 马鞍列岛海域虾类优势种密度对总密度的贡献夏季,虾类生物量和丰度与所有优势种相关,其中葛氏长臂虾的贡献率最大,β值分别为0.76和0.95,远远超过其他种类。秋季,虾类生物量与葛氏长臂虾、口虾蛄、中华管鞭虾和哈氏仿对虾相关,生物量贡献率以口虾蛄最高,β值为0.56。丰度与所有优势种相关,丰度贡献率则以葛氏长臂虾最高,β值为0.61。冬季,虾类生物量与葛氏长臂虾、细巧仿对虾、水母虾、细螯虾、口虾蛄和日本鼓虾相关,生物量贡献率以葛氏长臂虾最高,β值为0.88。虾类丰度与葛氏长臂虾、细巧仿对虾、水母虾和细螯虾相关,丰度贡献率以细巧仿对虾最高,β值为0.52,其次为葛氏长臂虾,β值为0.41。 春季,虾类生物量与葛氏长臂虾、细巧仿对虾、口虾蛄、水母虾、日本鼓虾和鲜明鼓虾相关,生物量贡献率以细巧仿对虾最高,β值为0.52,其次为口虾蛄和水母虾,β值均为0.31。虾类丰度与所有优势种相关,以水母虾的贡献率最高,β值为0.72,其次为细巧仿对虾,β值为0.49。
2.4 马鞍列岛海域虾类资源密度变化马鞍列岛海域夏季采获虾类30 515 ind,重量为58.97 kg;秋季采获虾类5 958 ind,重量为11.45kg;冬季采获虾类10 132 ind,重量为8.75 kg;春季采获虾类18 499 ind,重量为13.37 kg。调查海域全年虾类资源密度变化情况如图2所示,虾类年平均生物量和丰度分别为95.5 kg/km2和75.6×103 ind/km2,生物量和丰度均以夏季最高(206.7 kg/km2 和117.4×103 ind/km2),秋季最低(46.9 kg/km2和28.7×103 ind/km2)。生物量和丰度的最大值分别为最小值的4.4和4.1倍。

|
图2 马鞍列岛海域虾类资源密度变化 Fig.2 Seasonal variations of the resource density of shrimps in Ma′an Archipelago area |
单因素方差分析检验表明,虾类生物量夏季与秋、冬和春季均呈极显著差异(P=0.00、0.00、0.00<0.01);丰度夏季与秋季呈极显著差异(P=0.00<0.01),秋季与冬季呈显著差异(P=0.02<0.05),与春季呈极显著差异(P=0.00<0.01)。

|
表3 马鞍列岛海域虾类优势种密度对总密度的贡献 Tab. 3 Contribution of dominant species to total resource density of shrimps in Ma′an Archipelago area |
马鞍列岛海域种类丰富度(D)变化范围为0.69~2.08,最高值出现在夏季,最低值出现在春季;多样性指数(H′)变化范围为0.17~1.90,季节变动趋势与D值一致;均匀度指数(J′)变化范围为0.10~0.92,最高值出现在夏、秋季,最低值出现在春季。比较多样性指数均值(表2)可得:种类丰富度(D)、多样性指数(H′)和均匀度指数(J′)均呈现出秋季>冬季>夏季>春季的趋势。
单因素方差分析显著性检验结果表明,D值夏季与秋季、秋季与春季均呈极显著差异(P<0.01);H′值夏季与秋季和夏、秋、冬季与春季均呈显著性差异(P<0.05);J′值检验结果与H′值一致。
3 讨论 3.1 马鞍列岛海域虾类组成特征马鞍列岛海域全年出现虾类20种,从生态类群方面看[5]有60%为广温广盐种类。已有研究表明[5],东海30°30′N以北海域虾类种类组成主要为葛氏长臂虾、细巧仿对虾和中华管鞭虾等广温广盐种类。同步测定的环境数据显示,马鞍列岛海域表层水温变化范围为19.8~27.4℃,表层盐度变化范围为29.1~33.9,底层水温变化范围为20.2~24.7 ℃,底层盐度变化范围为30.0~35.8,水温和盐度年间变化范围较大,因此更适宜耐受性较强的广温广盐种类生存。马鞍列岛海域受长江冲淡水等沿岸低盐水影响[16],细螯虾、鲜明鼓虾和鞭腕虾等广温低盐种类广泛分布于马鞍列岛海域。葛氏长臂虾、细巧仿对虾、口虾蛄、哈氏仿对虾、日本鼓虾和水母虾6种广温广盐种类与鲜明鼓虾、细螯虾和鞭腕虾3种广温低盐种类在全年均有出现,占马鞍列岛海域虾类总种类数的45%,说明调查海域虾类种类组成以广温广盐种类占主要地位,广温低盐种类次之。水文环境的季节性变化对虾类种类组成影响很大[23],马鞍列岛海域水文环境受诸多因素影响,季节变化复杂,是导致其虾类组成生态类型多元化的主要原因。

|
表4 马鞍列岛海域虾类多样性指数 Tab. 4 The diversity indices of shrimps in Ma′an Archipelago area |
马鞍列岛海域虾类群落以葛氏长臂虾、细巧仿对虾和口虾蛄等广温广盐种类占主导地位。从马鞍列岛海域虾类优势种组成看,葛氏长臂虾为夏、秋和冬季虾类绝对优势种,细巧仿对虾为春季虾类绝对优势种,二者均属广温广盐生态类型。以水母虾为代表的广温低盐种类在冬春季相对重要性值明显提高,但由于其体型较小,生物量优势不明显(表2),因此仍以葛氏长臂虾、细巧仿对虾和口虾蛄等广温广盐种类为主要优势种。葛氏长臂虾和细巧仿对虾等虾类在马鞍列岛海域附近的中街山水域[10]以及舟山渔场[24]均为优势种,可见这些种类在近岸群岛水域具有较高的优势度。值得一提的是,马鞍列岛海域虾类不同种类适应岛礁环境的能力或者利用程度不同,所以其优势度有差别;而同一种类对不同季节岛礁生境的利用方式有别,所以集群性也有一定的季节变化,就造成了其在某些季节成为优势种而某些季节却非优势种的情况。
种类数上,马鞍列岛海域虾类总种类数与中街山列岛海域[10] 和椒江口海域[8]相差不大,约占舟山渔场及邻近海域[26]的57.1%,虾类组成均以广温广盐生态类群占主要地位。优势种数上,研究海域虾类总优势种数和各季节优势种数均高于中街山海域,但相比东海中北部海域[7]仍比较低。这一方面可能与调查海域面积、网具差异等有关[27](研究海域和东海中北部海域采用的为桁杆虾拖网,中街山海域采用的为单拖底拖网),马鞍列岛海域面积是中街山海域的2.7倍,其虾类种类数除夏季比中街山水域低,其他季节均比较高;另一方面与种类对环境的适应能力也有一定关系,除两个海域的共有优势种外,在研究海域哈氏仿对虾、鞭腕虾、水母虾、细螯虾、中国毛虾、鲜明鼓虾和脊腹褐虾在某些季节的生物量和丰度比例均低于10%,但由于其出现频率较高(表2),仍具有一定的优势地位,且这些种类都是r选择虾类,本身具备环境适应条件下大量繁殖的根基,由此形成了马鞍列岛海域虾类优势种组成相对丰富的特征。
春季,椒江口海域虾类优势种组成以中国毛虾和鲜明鼓虾等广温低盐种类占主导地位,研究海域水母虾和日本鼓虾等广温低盐种类的相对重要性值虽有所增加,但仍以广温广盐种类占主导地位。这主要是由于春季河口径流量增大,椒江口海域受沿岸低盐水的影响作用远比接近外海的研究海域明显,适宜的环境条件和丰富的营养物质[29]为鲜明鼓虾和细螯虾等广温低盐种类的繁殖以及优势地位的提高提供了良好的基础。秋季,台湾暖流携带的高温高盐水系成为影响两个海域的主要因子,其优势种组成则均以广温广盐种类占据绝对优势地位。由此可见,椒江口海域优势种组成相比研究海域在种类组成和生态类群方面的季节变化层次更丰富,但群落的主要特征随优势种的变化而发生相应的改变,优势种频繁更替不利于群落结构的稳定性[30]。另外,群落稳定性与物种多样性变化成反比关系[12],研究海域虾类群落多样性处于较低水平,而椒江口海域具备单个季节群落多样性水平较高的特点(表5),可见研究海域虾类群落比椒江口海域更趋于稳定状态。

|
表5 虾类种类数和优势种数比较 Tab. 5 Comparision of species and dominant species of shrimp |
葛氏长臂虾在夏、秋和冬季的相对重要性指数均高于其他种类,其各季节对虾类资源密度的贡献率也比较高,如葛氏长臂虾在夏、冬季对虾类生物量贡献率最高,夏、秋季对虾类丰度贡献率最高,可见,马鞍列岛海域虾类资源密度变化主要由葛氏长臂虾引起。细巧仿对虾和口虾蛄在某些季节对虾类资源密度也有较高的贡献率,如细巧仿对虾在春季对虾类生物量贡献率和冬季对虾类丰度贡献率均高于其他种类;口虾蛄在秋季对虾类生物量贡献率最高。葛氏长臂虾、细巧仿对虾和口虾蛄为虾类全年优势种,是浙江沿海最常见的虾类种类之一,因此广泛分布于马鞍列岛海域,夏秋季的出现频率更是在92%以上(表2)。这3个种类在不同季节对虾类生物量和丰度都有较大的贡献,可见其对温盐变化幅度较大的马鞍列岛海域具有广泛的适应性,因而能够成为马鞍列岛海域最重要的经济虾类,是影响调查海域虾类资源密度变化的主要种类。
3.4 马鞍列岛海域虾类群落多样性特征马鞍列岛海域虾类多样性存在显著性季节变化。多样性指数(H′)综合反映了虾类种类数和个体分配均匀性[9]。丰富度指数(D)、多样性指数(H′)和均匀度指数(J′)均以秋季最高,冬季次之,春季最低,丰富度指数(D)主要与种类数有关[28],多样性指数与种类数的变化趋势并不完全一致。这主要是由于葛氏长臂虾、细巧仿对虾、口虾蛄或水母虾等虾类的爆发式繁殖,导致其个体分配不均匀。如,夏季葛氏长臂虾的丰度比例为55.4%,形成单一丰度优势种格局。秋季虾类种类数最多,且除葛氏长臂虾外各优势种丰度比例相差不大,个体分配较均匀(表2),因此秋季的虾类多样性值最高。由此可见,种类间的个体均匀性是影响马鞍列岛海域虾类多样性变化的主要因素。与东海中北部海域[7]虾类群落多样性相比,马鞍列岛海域虾类群落丰富度指数(D)均值高于东海中北部海域,多样性指数(H′)均值则较东海中北部海域偏低,可见马鞍列岛海域虾类个体种间分配不均匀,是导致其多样性指数(H′)相对较低的主要原因,这从侧面说明葛氏长臂虾、细巧仿对虾和口虾蛄等个别虾类对近岸岛礁水域表现出强烈的依赖性。如,马鞍列岛海域(122°35′-l22°51′E;30°41′-30°52′N)葛氏长臂虾作为夏、秋和冬季的绝对优势种,其年平均生物量(28kg/h)为东海中部海域[6] (122°-l27°E;28°00′-31°00′N)(0.5 kg/h)的56倍。另外,与瓯江口、椒江口和东海中部海域相比(表6),马鞍列岛海域虾类资源除秋季生物量偏低,其他季节均处于较高水平,且其年平均生物量为东海中部海域的8.7倍。由此可见,环境条件较为复杂的马鞍列岛海域对虾类种群量的保障及多样性的维持起着极其重要的作用。
上海海洋大学邓明星、王坤和王家启等在现场采样过程中做了大量工作,嵊泗县渔民毛文勇协助本课题组成员完成了调查任务,沈天跃、席晓晴和曾旭等在实验中提供了大量帮助,谨致谢忱。
| [1] | GONZÁLEZ-GORDILLO J I, RODRÍGUEZ A. Comparative seasonal and spatial distribution of decapod larvae assemblages in three coastal zones off the south-western Iberian peninsula[J]. Acta Oecologica, 2003, 24(s1): S219-S233. |
| [2] | WENKAMP S, FISCHER P. The impact of coastal defence structures (tetrapods) on decapod crustaceans in the southern North Sea[J]. Marine Environmental Research, 2013, 92(12): 52-60. |
| [3] | MURA M, CAU A. Community structure of the decapod crustaceans in the middle bathyal zone of the Sardinian channel[J]. Crustaceana, 1994, 67(3): 259-266. |
| [4] | COSTA R C, FRANSOZO A, CASTILHO A L, et al. Annual, seasonal and spatial variation of abundance of the shrimp Artemesia longinaris (Decapoda: Penaeoidea) in south-eastern Brazil[J]. Journal of the Marine Biological Association of the United Kingdom, 2005, 85(1): 107-112. |
| [5] | 宋海棠, 俞存根, 薛利建, 等. 东海经济虾蟹类[M]. 北京: 海洋出版社, 2006. SONG H T, YU C G, XUE L J. Econmic Shrimp and Crab in East Sea[M]. Beijing: China Oceanic Press, 2006. |
| [6] | 宋海棠, 姚光展, 俞存根. 东海虾类的种类组成和数量分布[J]. 海洋学报, 2003, 25 (s1):171-179. SONG H T, YAO G Z, YU C G. Species composition and quantitative distribution of shrimps in the East China Sea[J]. Acta Oceanologica Sinica, 2003, 25(s1):171-179. |
| [7] | 陈小庆, 俞存根, 宋海棠, 等. 东海中北部海域虾类群聚结构特征及空间分布[J]. 海洋学研究, 2010, 28(4): 50-58. CHEN X Q, YU C G, SONG H T, et al. Structural characteristics and spatial distribution of the shrimp population in the mid-north East China Sea[J]. Journal of Marine Sciences, 2010, 28(4): 50-58. |
| [8] | 齐海明, 孙岳, 徐兆礼, 等. 椒江口海域春秋季虾类群落结构及其影响因素[J]. 应用生态学报, 2013, 24(12): 3546-3552. QI H M, SUN Y, XU Z L, et al. Shrimp community structure and its influential factors in the Jiaojiang river estuary during spring and autumn[J]. Chinese Journal of Applied Ecology, 2013, 24(12): 3546-3552. |
| [9] | 徐兆礼, 沈盎绿, 李新正. 瓯江口海域夏、秋季口足目和十足目虾类分布特征[J]. 中国水产科学, 2009, 16(1): 104-112. XU Z L, SHEN A L, LI X Z. Distribution of hoplocarida and decapoda shrimps in Oujiang estuary, East China Sea[J]. Journal of Fishery Sciences of China, 2009, 16(1): 104-112. |
| [10] | 谢汉阳, 朱文斌, 徐开达, 等. 中街山水域虾类组成及其群落多样性[J]. 广东海洋大学学报, 2012, 32(4): 1-7. XIE H Y, ZHU W B, XU K D, et al. Composition and community diversity of shrimps in the protected Zhongjieshan waters[J]. Journal of Guangdong Ocean University, 2012, 32(4): 1-7. |
| [11] | 晁文春, 何贤保, 苗振清, 等. 春夏季南麂列岛海域甲壳类种类组成及分布特征[J]. 浙江海洋学院学报(自然科学版), 2013, 32(3): 214-221. CHAO W C, HE X B, MIAO Z Q, et al. The species composition and diversity of crustaceans in the waters of Nanji islands[J]. Journal of Zhejiang Ocean University (Natural Science), 2013, 32(3): 214-221. |
| [12] | 汪振华, 王凯, 赵静, 等. 枸杞岛潮下带沙地生境鱼类群落结构和季节变化[J]. 应用生态学报, 2011, 22(5): 1332-1342. WANG Z H, WANG K, ZHAO J, et al. Fish community structure and its seasonal change in subtidal sandy beach habitat off southern Gouqi island[J]. Chinese Journal of Applied Ecology, 2011, 22(5): 1332-1342. |
| [13] | 金敬林, 蔡丽萍, 吴盈子. 马鞍列岛海洋特别保护区岩相潮间带底栖生物初步研究[J]. 海洋开发与管理, 2012(11): 80-84. JIN J L, CAI L P, WU Y Z. Prelininary investigation on the benthos of rocky intertidal zone in the Protected Ma'an Archipelago area [J]. Ocean Development and Management, 2012(11): 80-84. |
| [14] | 罗文俊, 章守宇. 马鞍列岛海域浮游植物优势种粒径组成及影响因子分析[J]. 上海海洋大学学报, 2009, 18(5): 571-578. LUO W J, ZHANG S Y. Analysis of equivalent spherical diameter composition and its influencing factors of dominant species of phytoplankton in Ma'an Archipelago[J]. Journal of Shanghai Ocean University, 2009, 18(5): 571-578. |
| [15] | 张启龙, 王凡, 赵卫红, 等. 舟山渔场及其邻近海域水团的季节特征[J]. 海洋学报, 2007, 29(5): 1-9. ZHANG Q L, WANG F, ZHAO W H, et al. Seasonal characteristics in the water masses in Zhoushan fishing ground and adjacent region[J]. Acta Oceanologica Sinica, 2007, 29(5): 1-9. |
| [16] | 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995. ZHAN B Y. Fish Stock Assessment[M]. Beijing: Chinese Agricultural Press, 1995. |
| [17] | 杨吝, 张旭丰, 谭永光, 等. 南海区拖网网囊最小网目尺寸选择性研究[J]. 中国水产科学, 2003, 10(4): 325-332. YANG L, ZHANG X F, TAN Y G, et al. Selectivity study on minimum mesh size of bottom trawl codend in South China Sea[J]. Journal of Fishery Sciences of China, 2003, 10(4): 325-332. |
| [18] | PINKAS L, OLIPHANT M S, IVRERSON I L K. Food habits of albacore, Bluefin tuna, and bonito in California waters[J]. Fish Bulletin, 1971, 152: 1-105. |
| [19] | MARGALEF R. Perspectives in Ecological Theory[M]. Chicago: University Chicago Press, 1968: 111-111. |
| [20] | SHANNON E C, WEAVER W. The Mathematical Theory of Communication [M]. Urbana: University of Illinois Press, 1948. |
| [21] | PIELOU E C. Species-diversity and pattern-diversity in the study of ecological succession[J]. Journal of theoretical biology, 1966, 10(2): 370-383. |
| [22] | CHRISTENSEN R. Analysis of Variance, Design, and Regression: Applied Statistical Methods [M]. New York: Chapman and Hall, 1996: 75-98. |
| [23] | 徐捷, 陈佳杰, 徐兆礼. 吕泗渔场沿岸海域春夏季虾类群落特征[J]. 水产学报, 2014, 38(8): 1097-1105. XU J, CHEN J J, XU Z L. Shrimp community structure in the Lvsi fishing ground during spring and summer[J]. Journal of Fisheries of China, 2014, 38(8): 1097-1105. |
| [24] | 王琳, 卢占晖, 苗振清, 等. 2012年春季舟山渔场虾类种类组成研究[J]. 浙江海洋学院学报(自然科学版), 2013, 32(1): 67-70. WANG L, LU Z H, MIAO Z Q, et al. The species composition of shrimp communities in spring in the Zhoushan fishing ground of 2012[J]. Journal of Zhejiang Ocean University (Natural Science), 2013, 32(1): 67-70. |
| [25] | 柏育材, 徐兆礼. 舟山渔场衢山岛海域春夏季十足目和口足目的分布特征[J]. 上海海洋大学学报, 2011, 20(1): 96-101. BAI Y C, XU Z L. Distribution of decapoda and hoplocarida shrimps near the sea area of Qushan Island during spring and summer[J]. Journal of Shanghai Ocean University, 2011, 20(1): 96-101. |
| [26] | 张少虎. 舟山渔场及邻近海域虾类种类组成与多样性分析[Z]. ZHANG S H. Analysis of species composition and diversity of shrimps in Zhoushan fishing ground and its adjacent waters[Z]. |
| [27] | 张洪亮, 张龙, 陈峰, 等. 浙江南部近岸海域春季蟹类群落结构特征[J]. 中国水产科学, 2013, 20(5): 1050-1056. ZHANG H L, ZHANG L, CHEN F, et al. Analysis of crab community characteristics in South Zhejiang coastal areas during spring[J]. Journal of Fishery Sciences of China, 2013, 20(5): 1050-1056. |
| [28] | 赵蒙蒙, 徐兆礼. 三门湾海域冬夏季口足目和十足目虾类的种类组成、时空分布及多样性分析[J]. 动物学杂志, 2011, 46(3): 11-18. ZHAO M M, XU Z L. Species composition, temporal and spatial distributions and diversity of hoplocarida and decapoda shrimps in summer and winter in Sanmen Bay[J]. Chinese Journal of Zoology, 2011, 46(3): 11-18. |
| [29] | 齐海明, 徐兆礼. 椒江口海域春秋季虾类资源分布与环境的关系[J]. 生态学杂志, 2013, 32(8): 2062-2069. QI H M, XU Z L. Shrimp resource distribution and its relationships with environment in the Jiaojiang Estuary of East China in spring and autumn[J]. Chinese Journal of Ecology, 2013, 32(8): 2062-2069. |
| [30] | 沈国英, 施并章. 海洋生态学[M]. 2版. 北京: 科学出版社, 2002. SHENG G Y, SHI B Z. Marine Ecology[M]. 2nd ed. Beijing: Science Press, 2002. |