 2016, Vol. 35
2016, Vol. 35扩展功能
文章信息
- 解崇友, 倪露芸, 吴迪, 王永明, 王志坚, 李斌
- XIE Chongyou, NI Luyun, WU Di, WANG Yongming, WANG Zhijian, LI Bin
- 岷江眉山段四川华鳊年龄与生长
- Age and Growth of Sinibrama taeniatus in Meishan Section of Minjiang River
- 四川动物, 2016, 35(1): 78-83
- Sichuan Journal of Zoology, 2016, 35(1): 78-83
- 10.11984/j.issn.1000-7083.20150141
-
文章历史
- 收稿日期: 2015-04-20
- 接受日期: 2015-07-24






2. 水产科学重庆市市级重点实验室, 西南大学生命科学学院, 重庆 400715
2. Key Laboratory of Aquatic Science of Chongqing, School of Life Sciences, Southwest University, Chongqing 400715, China
长江上游是我国生物多样性最丰富的地区之一,也是全球生物多样性热点之一(朱万泽等,2009)。鱼类资源是长江上游生物多样性的重要组成部分,已有数据显示长江上游鱼类种类260多种,其中特有鱼类118种(危起伟,2012)。四川华鳊Sinibrama taeniatus隶属鲤形目Cypriniformes鲤科Cyprinidae鮊亚科Cultrinae,俗称墨线鱼,主要分布于长江上游岷江、大渡河等水域,是我国长江上游特有小型经济鱼类(丁瑞华,1994)。近年来受采砂、水利工程建设、环境污染和酷渔滥捕等多种因素影响,四川华鳊资源量持续下降(骆辉煌等,2014)。因此有必要开展有关四川华鳊种质资源保护方面的研究工作。
鱼类年龄和生长规律是评价自然水域鱼类资源变动趋势的依据(邓中遴等,1981)。通过年龄与生长规律的研究,不仅可以掌握鱼类的生长特性(李文静等,2007),而且可以通过组建生长方程获得生长系数、拐点年龄等生长参数(颜云榕等,2011),可为鱼类种群动态(Gao et al.,2009)、渔业资源管理(Booth & Buxton,1997;Sun et al. ,2005;Govender et al.,2006)等提供基础数据。目前有关四川华鳊的研究仅见于分类学方面(丁瑞华,1994;谢仲桂等,2005)。本研究拟以长江上游岷江眉山段四川华鳊为实验材料,采用鳞片为年龄鉴定材料对其年龄与生长规律进行研究,旨在积累其生物学基础信息,为长江上游特有鱼类——四川华鳊资源的评估、保护与合理开发提供科学依据。
1 材料方法 1.1 标本采集与处理2013年3月至2014年1月在长江一级支流岷江眉山段采集四川华鳊样本共580尾,总质量6.053 kg,其中具有鳞片年龄信息的样本500尾。四川华鳊样本为沿江渔民用定置刺网、地笼等网具所捕获,有较强的随机性和代表性。样品采集后在鲜活状态下对其体长(L,mm)、体质量(W,g)等常规生物学特征进行测量,体长精确到1 mm,体质量精确到0.1 g。对样品进行现场解剖确定雌雄,并取其脊椎骨保存备用。同时取背鳍棘基部和侧线之间的鳞片8~10枚,立即放入5 mL离心管中加入纯净水保存带回实验室待用。
鳞片经水浸泡一段时间待表层上皮组织和色素细胞降解后,于体视镜(MOTIC SMZ-168 SERIES)下选取5~6枚形态完整、年轮清晰、轮纹连续的鳞片,逐一拍照(Moticam 2506),然后鉴定年龄,用Motic Images Advanced 3.2 软件测量轮径和鳞径,精确到0.01 μm。轮径是鳞焦到前区各年轮的距离,鳞径是鳞焦到前区轮纹边缘的距离。
1.2 数据分析
计算鳞片的边缘增长率(marginal growth increment,MGI),拟合体长与体质量、鳞径的回归方程。鳞片边缘增长率计算公式 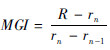 ;体长与体质量的关系表达式为W=aLb;体长与鳞径的关系表达式为L=a+bR,式中R为鳞径(mm),rn为鳞焦到最外一圈年轮轮纹的距离(mm),rn-1为鳞焦到次外圈年轮轮纹的距离(mm),a、b为系数。本研究中数据处理和分析在Excel 2003和SPSS 17.0中完成,图形处理在Photoshop CS5中完成。
;体长与体质量的关系表达式为W=aLb;体长与鳞径的关系表达式为L=a+bR,式中R为鳞径(mm),rn为鳞焦到最外一圈年轮轮纹的距离(mm),rn-1为鳞焦到次外圈年轮轮纹的距离(mm),a、b为系数。本研究中数据处理和分析在Excel 2003和SPSS 17.0中完成,图形处理在Photoshop CS5中完成。
四川华鳊的鳞片为圆鳞,较小且薄,易脱落。鳞焦偏于前区,形状似椭圆形和六边形,前端圆盾,后端呈扇形(图 1:a)。前区(基区)包藏在鳞囊内,绝大部分年轮标志为疏密型(图 1:b),宽阔环片(疏带)和狭窄环片(密带)区别明显。后区(顶区)部分露在鳞囊外,具有辐射的放射沟,环纹被放射沟所截断。侧区年龄特征不明显,少部分能观测到切割现象(图 1:c)。因此,后续研究中鳞径和轮径的测量都是以鳞焦到前区之间的距离为准。鳞片中也发现有再生鳞存在的现象(图 1:d)。

|
|
图 1 四川华鳊的鳞片信息
Fig. 1 The scale information of Sinibrama taeniatus
a. 完整形态, b. 疏密年轮, c. 切割年轮, d. 再生鳞。 a. Integral structure, b. Loose-close type, c. Incision type, d. Regeneration of scale. |
年龄鉴定显示,绝大多数四川华鳊样本的年龄介于1+ ~2+之间(1+龄数量多于2+龄),因此按月份分组计算了1+龄和2+龄鳞片的边缘增长率。图 2显示1+龄和2+龄鳞片MGI峰值均出现在12月,对应值分别为1.40和1.69。

|
| 图 2 四川华鳊鳞片边缘增长率的月变化 Fig. 2 Monthly changes of scale marginal growth increment (MGI) of Sinibrama taeniatus |
对580尾四川华鳊样本进行统计,其中最小体长59 mm,最大体长113 mm,平均体长82.34 mm,主要分布于71~100 mm,占总数的88.80%;最小体质量3.8 g,最大体质量为28.5 g,平均体质量10.58 g,主要分布在6.0~20.0 g,占总数的88.30%。在有 鳞片年龄信息的500尾样品中,年龄组成中最大个体为3+龄,以1+、2+龄为优势类群,占已确定年龄 的96.07%。样本中雄性个体329尾,雌性个体211尾,肉眼未能分辨性别有40尾,种群中雄、雌性比例达到1.57∶ 1。
2.4 体长与体质量的关系鱼类体长和体质量之间存在一定的相关性,同种体长的鱼,体质量越大说明其营养状况和生长环境越佳(殷名称,1995)。本研究将四川华鳊雌鱼和雄鱼的体长与体质量之间的相关性分别进行了拟合(图 3)。在多种拟合函数关系中,幂函数模型的相关指数是最高的,故选择了幂函数模型。四川华鳊雌鱼样本体长与体质量拟合的方程为W=2×10-5L3.03(R2=0.89,n=211),雄鱼样本方程为W=3×10-5L2.91(R2=0.86,n=329)。经方差检验,雌、雄鱼样本间差异无统计学意义(P>0.05),所以公式统一为W=2.4×10-5L2.94。

|
| 图 3 四川华鳊体长与体质量的关系 Fig. 3 Relationship between standard length and total weight of Sinibrama taeniatus |
选用线性、对数、乘幂、多项式和指数5种回归模型对四川华鳊体长和鳞径的数据进行拟合,结果表明线性方程L=a+bR拟合度最高(图 4)。拟合方程分别为,雌鱼样本L=25.82R+67.65(R2=0.33),雄鱼样本L=16.19R+71.18(R2=0.15);经方差检验雌、雄鱼样本之间差异无统计学意义(P>0.05),公式统一为L=25.45R+65.73。

|
| 图 4 四川华鳊体长与鳞径的关系 Fig. 4 Relationship between standard length and scale radius of Sinibrama taeniatus |
四川华鳊的体长和鳞径呈线性相关,且直线不通过坐标轴原点,故可采用Rose Lee退算各龄组的体长(殷名称,1995)。将退算体长(表 1)和实测平 均体长进行配对的t检验,结果表明退算体长和实测体长间差异无统计学意义。
| 年龄组 Age group | 实测平均体长 Observed mean length/mm | 各龄鱼退算体长 Back calculated length/mm | 差值 Difference value/mm | 样本数 Number | ||
| L1 | L2 | L3 | ||||
| 0+ | 71.6 | 3.06 | 18 | |||
| 1+ | 81.4 | 75.10 | -0.65 | 401 | ||
| 2+ | 87.8 | 71.91 | 80.74 | 5.43 | 64 | |
| 3+ | 113.0 | 74.19 | 81.52 | 93.23 | 1 | |
| 按标本数平均 Mean based on the sample number | 74.66 | 80.75 | 93.23 | |||
四川华鳊体长与体质量关系式中W=2.4×10-5L2.94,b=2.94,趋近于3,表明该鱼属于等速生长类型(Santos et al.,2002)。由于四川华鳊总体上属于等速生长类型,因此其生长特征可用Von Bertalanffy方程进行描述。体长和体质量的生长方程分 别为:Lt=L∞,Wt=W∞ b。 式中,Lt、Wt分别表示年龄为t时的体长和体质量;L∞、W∞分别表示渐进体长和渐进体质量;K为生长曲线的曲率参数;t0为理论生长起始时间,为假定的理论常数。依据四川华鳊渔获物各龄的退算体长,用最小二乘法算得生长方程的各参数L∞=139.57 mm,K=0.15,t0=-3.16,W∞=47.97。将各参数代入Von Bertalanffy方程,得到体长和体质量的生长方程为Lt=139.57×,Wt=47.97× 2.94。图 5显示了四川华鳊体长生长曲线无明显拐点,而是趋向于渐进体长。体质量生长曲线呈非对称的“S”形,前期呈现加速趋势,到拐点处生长速度达到最大,之后逐渐下降并趋向于渐进体质量。

|
| 图 5 四川华鳊体长和体质量生长曲线 Fig. 5 Growth curves of standard length and total weight of Sinibrama taeniatus |
对四川华鳊的体长和体质量生长方程进行一阶和二阶求导,获得体长和体质量的生长速度和生长加速度方程。体长生长速度方程:
dl/dt=139.57×0.15×e-0.15(t+3.16)
体长生长加速度方程:
d2l/d2t=-139.57×0.152×e-0.15(t+3.16)
体质量生长速度方程:
dw/dt=2.94×47.97×0.15×e-0.15(t+3.16) 1.94
体质量生长加速度方程:
d2w/d2t=2.94×47.97×0.152×e-0.15(t+3.157) 0.94
岷江眉山段四川华鳊体长生长速度和加速度均无明显的拐点,体长生长速度随着年龄的增长呈递减趋势,且递减趋势减慢,缓慢趋于0(图 6)。体长生长加速度先呈递增趋势,然后逐渐变缓,但加速度的增加始终小于0,说明了四川华鳊幼龄时生长速度较快,随着年龄的增大,体长生长开始变缓。 图 7显示了四川华鳊体质量生长速度和加速度具有明显生长拐点,体质量生长速度最大时即生长拐点(i),拐点年龄ti=lnb/k+t0=4.09龄,对应的体长和体质量分别为Li=92.06 mm,Wi=14.13 g。生长拐点前体质量生长速度呈递增趋势,拐点后生长速度呈递减趋势。生长加速度逐渐减小,在生长拐点处加速度为0。

|
| 图 6 四川华鳊体长的生长速度和生长加速度随年龄的变化曲线 Fig. 6 The growth rate and growth accelaration curves of standard length of Sinibrama aeniatus |

|
| 图 7 四川华鳊体质量的生长速度和生长加速度随年龄的变化曲线 Fig. 7 The growth rate and growth accelaration curves of total weight of Sinibrama taeniatus |
准确获取鱼类种群年龄结构是评估鱼类资源状态、制定鱼类资源管理模式(Hilborn & Walters,1992)的重要依据。通常低估鱼群年龄结构会引起鱼类种群生长率和死亡率计算结果偏大,而高估则会导致鱼类种群资源的过度捕捞(Eklund et al.,2000;沈建忠等,2001)。鱼类的年龄鉴定一般采用鳞片、耳石、 鳍条和脊椎骨等材料进行(殷名称,1995)。邓中遴等(1981)指出在鱼类年龄鉴定的众多材料中,鳞片是最方便取材的,尤其适用于低龄且生长较迅速的小型鱼类。目前已经有多位学者采用鳞片对鮊亚科多种小型鱼类进行了年龄鉴定。刘国栋(2011)对大眼华鳊S. macrops,王俊(2012)对半 Hemiculterella sauvagei,孙宝柱(2010)对张氏 Hemiculter tchangi等均采用了鳞片作为年龄鉴定材料,并且得到较 为理想的研究结果。本研究主要采用鳞片 对四川华鳊的年龄进行了鉴定。四川华鳊鳞片年轮结构清晰,年龄特征为普通疏密型,副轮较少,生长中心较为明确,便于测量鳞径和轮径,能准确确定其年龄,同时也增加了与其他学者研究结果的可比性。
Hemiculterella sauvagei,孙宝柱(2010)对张氏 Hemiculter tchangi等均采用了鳞片作为年龄鉴定材料,并且得到较 为理想的研究结果。本研究主要采用鳞片 对四川华鳊的年龄进行了鉴定。四川华鳊鳞片年轮结构清晰,年龄特征为普通疏密型,副轮较少,生长中心较为明确,便于测量鳞径和轮径,能准确确定其年龄,同时也增加了与其他学者研究结果的可比性。
探讨鱼类体长与体质量之间的关联对渔业生产和研究种群数量变化规律有重要指导意义(薛超等,2013)。研究中四川华鳊生长系数K=0.15,Branstetter(1987)认为,K值大于0.10属于生长相对较快的鱼类。岷江眉山段位于长江上游生物多样性区域内,该区域温度适宜,水体生产力丰富,饵料资源也相对丰富,四川华鳊生长速度相对较快。
本研究将四川华鳊生长特性与同亚科的几种小型鱼类进行了比较(表 2)。结果发现四川华鳊(4.09龄)与同属的大眼华鳊(4.35龄)生长拐点较为接近,在这几种小型鱼类中属于生长潜力较大的种群,但其生长拐点所对应的体长、体质量指标在这几种鱼类中却是最低的(表 2);在580尾四川华鳊样本中,体长在71~100 mm范围内的个体占总数的88.8%,体质量在6.0~20.0 g 范围内的个体占88.3%。2~3龄的个体占96.07%(其中以2龄个体数量为主)。上述结果表明,目前分布在长江上游岷江眉山段四川华鳊种群已经出现了小型化趋势。
| 比较项目 Item | 四川华鳊 Sinibrama taeniatus | 大眼华鳊 S. macrops | 张氏 Hemiculterella tchangi | 半 H.sauvagei | H.leucisculus | |
| 生境 Habitat | 岷江 Minjiang River | 千岛湖 Thousand-island Lake | 长潭水库 Changtan Reservoir | 赤水河口 Chishui Estuary | 赤水河赤水市 Chishui County | 北江 Bei River |
| b | 2.9377 | 2.8985 | 3.0518 | 3.1450 | 2.8720 | 3.0736 |
| L∞/mm | 139.57 | 240.40 | 139.80 | 196.05 | 176.10 | 283.84 |
| W∞/g | 47.97 | 249.38 | 56.01 | 97.20 | 59.54 | 280.82 |
| Li | 92.06 | 157.40 | 109.10 | 133.50 | 114.78 | 191.50 |
| Wi | 14.13 | 73.13 | 16.64 | 29.13 | 17.41 | 83.77 |
| K | 0.15 | 0.15 | 0.68 | 0.36 | 0.26 | 0.18 |
| t0 | 3.16 | 2.85 | 0.33 | 1.26 | 1.13 | 1.35 |
| ti | 4.09 | 4.35 | 1.95 | 1.92 | 2.71 | 4.96 |
| 文献来源 | 本研究 | 刘国栋等,2011 | 陆清尔,1992 | 孙宝柱等,2010 | 王俊等,2012 | 李强等,2009 |
四川华鳊作为我国长江上游特有鱼类,加强对其研究和保护具有重大的科学价值和生态价值。保护工作可从以下几个方面开展:(1)加强对栖息环境的保护。(2)禁止非法捕捞。鉴于本研究中渔获物四川华鳊年龄小于拐点年龄,且低龄组占大部分,在理论推导上出现了个体小龄化现象,但缺乏时间序列上的对比,应加强渔业管理力度。首先严格执法,禁止每年的4—5月对渔业资源的偷捕活动(4—5月为四川华鳊的主要繁殖时间);其次取缔非法捕捞工具,如电捕、小网眼刺网,禁止使用网眼小于2 cm的捕捞工具。(3)科学研究、保护和开发。例如加强四川华鳊的人工繁殖、资源动态、种群特征及遗传特性方面的研究工作,积极开展四川华鳊资源增殖放流和生态恢复技术研究,补充和恢复四川华鳊自然种群数量。(4)观赏与保护。四川华鳊因体侧具乌黑的侧线,有一定的观赏价值,深受喜爱,将其作为观赏鱼打造,既能起到对四川华鳊这一长江上游特有鱼类资源的保护作用,又可获得一定的经济效益与社会效应。
| 邓中遴, 余志堂, 许蕴玕, 等. 1981. 汉江主要经济鱼类的年龄与生长[M]// 鱼类学论文集: 第一辑. 北京: 科学出版社: 97-111. |
| 丁瑞华. 1994. 四川省鱼类志[M]. 成都: 科学出版社. |
| 李强, 赵俊, 钟良明, 等. 2009. 北江生长研究[J]. 广州大学学报, 8(4): 38-40. |
| 李文静, 王剑伟, 谢从新, 等. 2007. 厚颌鲂的年龄结构与生长特性[J]. 中国水产科学, 14(2): 215-222. |
| 刘国栋, 何光喜, 刘其根, 等. 2011. 千岛湖大眼华鳊年龄、生长和繁殖的初步研究[J]. 上海海洋大学学报, 20(3): 382-391. |
| 陆清尔. 1992. 长潭水库大眼华鳊(Sinibrama macrops)年龄与生长的初步研究[J]. 浙江水产学院学报, 2(11): 116-122. |
| 骆辉煌, 杨青瑞, 李倩, 等. 2014. 长江上游珍稀特有鱼类保护区鱼类资源生境特征初步研究[J]. 淡水渔业, 44(6): 44-48. |
| 沈建忠, 曹文宣, 崔奕波. 2001. 用鳞片和耳石鉴定鲫年龄的比较研究[J]. 水生生物学报, 25: 462-466. |
| 孙宝柱, 李缙, 但胜国, 等. 2010. 张氏䱗的繁殖生物学特性[J]. 水生生物学报, 34(5): 998-1003. |
| 王俊, 王美荣, 但胜国, 等. 2012. 赤水河半䱗年龄与生长[J]. 四川动物, 31(5): 713-719. |
| 危起伟. 2012. 长江上游珍稀特有鱼类国家级自然保护区科学考察报告[M]. 北京: 科学出版社. |
| 谢仲桂, 谢从新, 张鹗. 2005. 我国华鳊鱼类形态差异及其物种有效性的研究[J]. 动物学研究, 24(5): 321-330. |
| 薛超, 危起伟, 孙庆亮, 等. 2013. 秦岭细鳞鲑的年龄与生长[J]. 中国水产科学, 20(4): 743-749. |
| 颜云榕, 侯刚, 卢伙胜, 等. 2011. 北部湾斑鳍白姑鱼的年龄与生长[J]. 中国水产科学, 18(1): 145-155. |
| 殷名称. 1995. 鱼类生态学[M]. 北京: 中国农业出版社. |
| 朱万泽, 范建容, 王玉宽, 等. 2009. 长江上游生物多样性保护重要性评价--以县域为评价单元[J]. 生态学报, 29(5): 2603-2611. |
| Booth AJ, Buxton CD. 1997. Management of the panga Pterogymnus laniarius (Pisces: Sparidae) on the Agulhas Bank, South Africa using per-recruit models[J]. Fisheries Research, 32: 1-11. |
| Branstetter S. 1987. Age and growth estimates for blacktip, Carcharhinus limbatus, and spinner, C. brevipinna, sharks from the northwestern Gulf of Mexico[J]. Copeia, (4): 964-974. |
| Eklund J, Parmanne R, Aneer G. 2000. Between-reader variation in heering otolith ages and effects on estimated population parameters[J]. Fisheries Research, 46: 147-154. |
| Gao X, Brosse S, Chen YB, et al. 2009. Effects of damming on population sustainability of Chinese sturgeon, Acipenser sinensis: evaluation of optimal conservation measure[J]. Enviromental Biology of Fishes, 86: 325-336. |
| Govender A, Al-Oufi H, Mcllwain JL, et al. 2006. A per-recruit assessment of the kingfish (Scomberomorus commerson) resource of Oman with an evaluation of the effectiveness of some management regulations[J]. Fisheries Research, 77: 239-247. |
| Hiborn R, Walters CJ. 1992. Quantitative fisheries stock assessment: Choice, dynamics, and uncertainty[M]. New York: Chapman and Hall: 570. |
| Santos MN, Gaspar MB, Vasconcelos P, et al. 2002. Wight-length relationships for 50 selected fish species of the Algare cosat (southern Portugal)[J]. Fisheries research, 59: 289-295. |
| Sun CL, Wang SP, Porch CE, et al. 2005. Sex-specific yield per-recruit and spawning stock biomass per-recruit for the swordfish, Xiphias gladius, in the waters around Taiwan[J]. Fisheries Research, 71: 61-69. |