 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 张守栋, 杨传华, 林恭华, 张同作, 苏建平
- ZHANG Shoudong, YANG Chuanhua, LIN Gonghua, ZHANG Tongzuo, SU Jianping
- 高寒草甸3种啮齿动物内脏器官及相关性比较
- Comparison of the Internal Organs and Correlation of Organ Weights and Body Weights of Three Rodents in Alpine Meadow
- 四川动物, 2015, 34: 424-428
- Sichuan Journal of Zoology, 2015, 34: 424-428
- 10.3969/j.issn.1000-7083.2015.03.020
-
文章历史
- 收稿日期:2014-09-08
- 接受日期:2014-12-05






2. 浙江省湖州市现代农业技术学校, 浙江湖州313023;
3. 中国科学院大学, 北京100049
2. Modern Agricultural Technical School of Huzhou, Huzhou, Zhejiang Province 313023, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
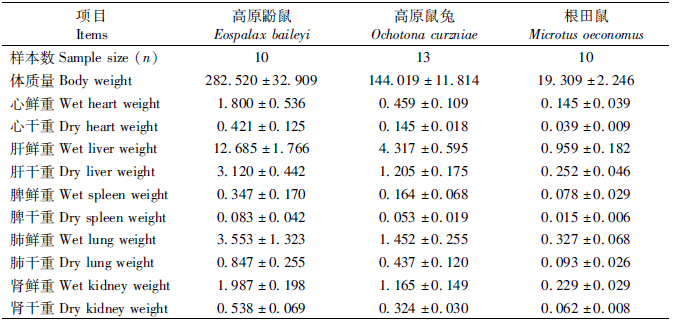
脏器指数是动物某脏器的质量与其体质量的比值,脏器质量和脏器指数是动物主要的生物学特性指标之一(詹纯列等,2001)。内脏器官是动物生理功能的物质基础,其质量及脏器指数往往是一定环境条件下生理功能的反映,与动物的生长发育、繁殖、季节及栖息环境变化有关(Pucek,1965;Schieck & Millar,1985;王淑卿等,1999;杜卫国等,1999;张美文等,2006)。比较内脏器官可以间接反映动物的环境适合度。国内外不少学者探讨了野外条件下鼠类脏器指数及其生态适应意义(Pucek,1965;Schieck & Millar,1985;王德华,孙儒泳,1995)。Pucek(1965)认为脏器指标是动物代谢功能的良好指标,高代谢率的小动物一般具有相对较大的心脏。叶润蓉和曹伊凡(1994)、赵建文等(1999)对高原鼠兔Ochotona curzoniae的内脏器官进行过解剖观察和质量测定。张世炎等(2001)对黄毛鼠Rattus lossea部分内脏器官的质量、相对质量(脏器指数)及含水量进行了测定,发现黄毛鼠内脏器官在年龄和性别间存在差异。胡忠军等(2002)对实验室封闭群第7、8代东方田鼠Microtus fortis calamorum头骨和脏器的形态学指标进行了测量,并探讨了各指标在雌雄间、年龄组间的差异及其与体质量的关系。杜卫国等(1999)研究了社鼠R. niviventer主要内脏器官的绝对质量、相对质量(脏器指数)和含水量的季节变化。
青藏高原地区气候为典型的大陆性高寒气候,其特点表现为冬季寒冷且漫长。高原鼢鼠Eospalax baileyi、高原鼠兔和根田鼠M. oeconomus是青藏高原优势植食性小哺乳动物(崔庆虎等,2005;王晓君等,2008)。3种草食哺乳动物是高寒草甸生境中最主要的鼠种,且常见到生境重叠的现象(王祖望等,1979;王玉山等,2001)。高原鼢鼠属啮齿目Rodentia鼹形鼠科Spalacidae凸颅鼢鼠属Eospalax(Norris et al.,2004;Zhou & Zhou,2008),高原鼠兔属兔形目Lagomorpha鼠兔科Ochotonidae鼠兔属Ochotona(冯祚建,郑昌琳,1985),根田鼠属啮齿目Rodentia仓鼠科Cricetidae田鼠属Microtus(李艳萍,2007)。高原鼢鼠是终生生活在完全封闭的地下洞道中的地下鼠,洞道生境中氧含量显著低于同地区大气氧含量,同时洞道中含高浓度的CO2。而高原鼠兔与根田鼠在地面和开放洞道中生活,其生境的氧含量相对较高,CO2含量很低。繁殖期鼠类能量需求会增加,繁殖期雌鼠的能量需要明显高于非繁殖雌鼠(Millar,1977)。
目前,国内外有关野外啮齿类动物内脏器官的研究做了大量工作。但通过对高寒草甸小型啮齿类动物内脏器官比较及脏器质量与体质量的相关性研究来探讨动物对环境的适应性未见报道。本文通过对青藏高原高寒草甸3种小型啮齿动物脏器指数及脏器质量与体质量相关性的比较,试图了解内脏器官与动物进化适应的关系,并为进一步探讨青藏高原高寒草甸中动物对环境的适应提供依据。 1 材料与方法 1.1 试验动物及自然概况
张克勤等(2008)认为啮齿类器官的性别差异特征随着种类不同而不同。为排除繁殖状态的影响,本研究所选用的动物均为雄性。2012年5月于青海省大通县东峡镇(海拔2994 m,地理位置37°03'N,101°47'E,年均气温2.1 ℃)捕获10只雄性高原鼢鼠;2012年7月于中国科学院海北高寒草甸生态系统定位站(平均海拔3200 m,地理位置37°29'~37°45'N,101°12'~101°23'E)捕获13只成年雄性高原鼠兔,10只雄性根田鼠。高原鼢鼠采集方法为地箭死捕法,高原鼠兔采集方法为绳套法,根田鼠采集方法为U形铁丝笼法。两地生境类型均为高寒草甸生态系统,土壤较湿润。由于受东南季风和西伯利亚高压的影响,该地区呈典型的大陆高寒气候,无明显四季之分,仅有冷暖二季。 1.2 器官组织的解剖
捕获动物用电子天平称重(精确到0.01 g),经冷冻处理带回实验室。试验在中国科学院西北高原生物研究所高原生物适应与进化重点实验室进行。试验参考杨传华等(2012)的方法。解剖时由腹部向上纵向剪开,小心分离并取出内脏器官,仔细剔除其附带的结缔组织和白色脂肪组织,并置于电子天平上称重(精确到0.01 g),获取心、肝、脾、肺、肾的鲜重。将上述样品在60 ℃条件下烘干至恒重,测定其干重。 1.3 数据统计分析
脏器指数的计算公式见杜卫国等(1997)的方法。即:脏器指数=该脏器鲜重/体质量×100%。数据利用SPSS 19.0进行处理。分析前,所有数据均用Kolmogorov-Smirnov检验其正态性,方差齐性用Levene检验,非正态分布的数据经转换标准化后使用。用单因素方差分析(One-Way ANOVA)及最小显著差数法(LSD)和Tamhanes T2多重比较检验各相关变量。用相关分析研究脏器质量与体质量的相关性。文中数据以平均值±标准差(Mean±SD)表示,P<0.05即认为差异有统计学意义。 2 结果 2.1 内脏器官质量及脏器指数的比较
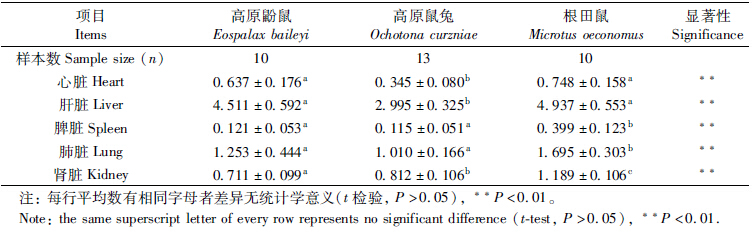
高寒草甸3种啮齿动物的体质量及各内脏器官质量在表 1中列出。3种啮齿动物的脏器指数比较结果见表 2。One-Way ANOVA结果显示,3种啮齿动物的体质量及内脏器官比值差异有高度统计学意义(P<0.01)。各动物间脏器指数的差异不尽相同。LSD和Tamhanes T2多重比较显示,高原鼢鼠与根田鼠的心脏及肝脏指数的差异无统计学意义(P>0.05),但明显大于高原鼠兔;根田鼠的脾脏及肺脏指数大于高原鼢鼠与高原鼠兔,且差异有高度统计学意义(P<0.01);肾脏指数的差异为根田鼠>高原鼠兔>高原鼢鼠,且差异有高度统计学意义(P<0.01);根田鼠的胃指数大于高原鼢鼠与高原鼠兔,且差异有高度统计学意义(P<0.01);高原鼠兔的小肠及大肠指数大于高原鼢鼠与根田鼠,且差异有高度统计学意义(P<0.01);盲肠指数的差异为高原鼠兔>高原鼢鼠>根田鼠,且差异有高度统计学意义(P<0.01)。
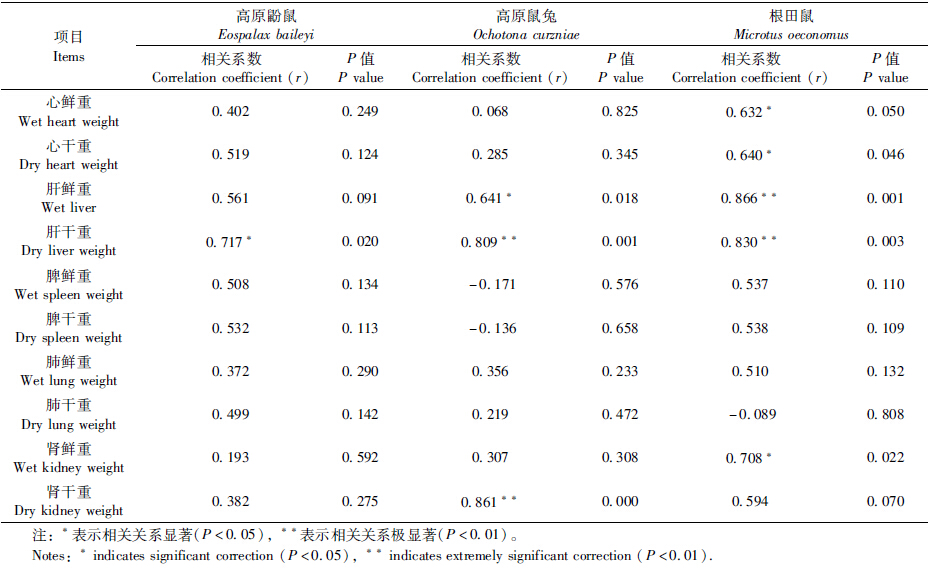
3种啮齿动物脏器质量与体质量的相关性见表 3。结果显示,3种啮齿动物肝干重与体质量的相关关系表现出一致的变化规律,均与体质量呈显著正相关关系。高原鼢鼠其他脏器质量与体质量之间无显著相关关系。高原鼠兔和根田鼠的肝鲜重与体质量之间有显著正相关关系。根田鼠的心鲜重和心干重与体质量显著正相关。高原鼠兔的肾干重与根田鼠的肾鲜重与体质量表现出显著的正相关关系。其他各器官与体质量之间无显著相关关系。
脏器质量和脏器指数是动物主要的生物学特性指标之一(詹纯列等,2001)。内脏器官作为动物生理功能的载体,其指数近似反映该脏器的生理指标(张世炎,麦海,2001)。因此,脏器指数可以代表动物的代谢功能,高代谢率的小动物一般具有相对较大的心脏(Pucek,1965)。心脏质量的增加有利于提高抵御严寒的能力(张志强,王德华,2009)。心脏是全身组织的供氧中枢和低氧代谢产物乳酸的主要清除器官之一(刘国富等,1985)。高原鼢鼠和高原鼠兔虽然同为高原动物,但是它们具体生境存在很大差异。高原鼢鼠终生营地下生活,低O2高CO2是其生境主要特点;而高原鼠兔为地面动物,与高原鼢鼠相比,其主要应对低氧胁迫(王晓君等,2008)。相对较大的心脏指数有利于高原鼢鼠清除低氧代谢产物。根田鼠个体小,相对体表面积较大,散失的热量相对较高,心脏指数的增大有利于其抵御严寒。一般而言,心脏指数是代谢功能的良好指标,高代谢率的小动物具相对较大的心脏(杜卫国等,1999)。根田鼠的最大代谢率高于高原鼠兔的最大代谢率(王玉山等,2001),高原鼢鼠的代谢率明显低于高原鼠兔(王祖望等,1979)。同样可以解释高原鼢鼠和根田鼠的心脏指数大于高原鼠兔。
肾脏有重吸收水分的功能,高原鼢鼠生活在密闭的洞道中,空气湿度较高,肾脏重吸收水分的能力相对低于地上动物。根田鼠相对大的体表面积需要相对较大的肾脏以降低水分丧失。根田鼠相对较大的肾脏指数可能是维持代谢和维持身体水分综合需要的进化结果。
栖居于高海拔地区的小哺乳动物受到低温和低氧双重胁迫,而两者对代谢率的影响则相反,低氧降低了肺承载氧的能力,这可能制约了氧耗的最大速率,也就限制了代谢产热(Rosenmann & Morrison,1974)。根田鼠较高的代谢率需要相对较大的肺来增加载氧能力。
肝脏是小哺乳动物体内一个重要的产热器官,静止状态下其产热可占总体产热的25%左右(Schmidt-Nielsen & Knut,1997)。地下活动鼠类主要通过挖掘土壤以获取食物,主要取食植物的根茎等器官(王德华,王祖望,2000)。Vleck(1979)曾对囊鼠Thomomys bottae的挖掘能量消耗做过测定,发现挖掘与在地表运动相同的距离,所消耗的能量比地表高出360~ 3400倍。高原鼢鼠在地下为获取食物所消耗的能量大,根田鼠为维持体温必须有相对较高的产热。因此高原鼢鼠与根田鼠的肝脏指数比高原鼠兔的高。3种啮齿动物肝干重均与体质量呈显著正相关关系。高原鼠兔和根田鼠的肝鲜重与体质量之间有显著正相关关系。可见3种动物的肝脏质量与体质量的相关关系表现出一致的变化规律,这说明肝脏可能受动物体型的影响较大,与动物的产热、散热及能量消耗密切相关。
根田鼠的脾脏指数较高原鼢鼠与高原鼠兔的高。脾脏是哺乳动物体内具有重要免疫功能的器官,它不仅是T细胞和B细胞定居的场所,而且是机体对血源性抗原产生免疫应答的主要场所(边疆晖等,2008)。脾脏不仅能合成免疫活性物质,而且其红髓中的巨噬细胞可清除血液中的外来抗原(边疆晖等,2008)。根田鼠属于地上动物,接触抗原风险较高原鼢鼠大,加上个体小,必须进化出相对较大的免疫器官来保证个体不会受到病原体感染。
关于脏器质量与体质量的相关性,3种动物的肝脏质量与体质量的相关关系表现出一致的变化规律。说明肝脏可能受动物体型限制,主要与动物的产热、散热及能量消耗密切相关。根田鼠的心脏质量与体质量显著正相关,而高原鼢鼠与高原鼠兔的心脏质量与体质量无显著相关关系。这说明体型较小的动物必须有一个大的心脏来满足自身血液循环及能量代谢的需求。高原鼠兔的肾干重与根田鼠的肾鲜重与体质量表现出显著的正相关关系,而高原鼢鼠的肾脏质量与体质量无显著相关关系。这可能与高原鼢鼠是地下鼠,洞道湿度高;高原鼠兔和根田鼠在地面活动,必须有与体型相适应的肾脏来重吸收水分,减少水分散失。3种动物的脾脏质量与体质量均无显著相关性,这可能由于脾脏是机体主要的免疫器官(边疆晖等,2008),动物生活的环境不同,所面临的外界病原体明显不同。脾脏必须根据生活环境所面临的免疫压力做出调整,而不是与体型大小相适应。
在器官水平上,青藏高原高寒草甸3种小型啮齿动物的脏器指数及脏器质量与体质量的相关性都呈现出不同程度的差异。这些差异体现出不同物种面对不同的生存环境做出的适应性进化。青藏高原环境苦寒严酷,高原生物要想在如此严酷的环境中生存、繁衍就必须做出形态、生理适应。而不同动物由于体型大小及所处环境差异等所做出的适应对策不同。不同物种脏器指数及脏器质量与体质量的相关性差异可能是物种对物质环境、能量环境、生物个体大小、外界病原体、能量代谢需求等多方面综合因素进化适应的结果。
| 边疆晖, 曹伊凡, 吴雁, 等. 2008. 青藏高原地区根田鼠脾脏重量及迟发性超敏反应的季节性变化[J]. 兽类学报, 28(3): 242-249. |
| 崔庆虎, 蒋志刚, 连新明, 等. 2005. 根田鼠栖息地选择的影响因素[J]. 兽类学报, 25(1): 45-51. |
| 杜卫国, 鲍毅新, 施利强, 等. 1999. 社鼠内脏器官重量和水分含量的季节变化[J]. 动物学杂志, 34(1): 23-25. |
| 杜卫国, 俞华英, 施利强, 等. 1997. 褐家鼠的身体及内脏器官重量和含水量的初步研究[J]. 中国媒介生物学及控制杂志, 8(3): 161-163. |
| 冯祚建, 郑昌琳. 1985. 中国鼠兔属(Ochotona)的研究——分类与分布[J]. 兽类学报, 5(4): 269-289. |
| 胡忠军, 王勇, 张美文, 等. 2002. 东方田鼠头骨和脏器的形态学指标[J]. 动物学杂志, 37 (4):21-26. |
| 李艳萍. 2007. 根田鼠雄性生殖腺的胚后发育[D]. 西安: 陕西师范大学. |
| 刘国富, 温得启, 胡晓梅. 1985. 高原鼠兔和高原鼢鼠乳酸脱氢酶同工酶的初步研究[J]. 兽类学报, 5(3): 223-228. |
| 王德华, 孙儒泳. 1995. 根田鼠消化道长度和重量的变化及其适应意义[J]. 兽类学报, 15(1): 53-59. |
| 王德华, 王祖望. 2000. 高寒地区高原鼢鼠消化道形态的季节变化[J]. 兽类学报, 20(4): 270-276. |
| 王淑卿, 张知彬, 张健旭, 等. 1999. 大仓鼠消化道长度和重量变化的初步研究[J]. 动物学杂志, 34(6): 17-21. |
| 王晓君, 魏登邦, 魏莲, 等. 2008. 高原鼢鼠和高原鼠兔肺细叶的结构特征[J]. 动物学报, 54(3): 531-539. |
| 王玉山, 王德华, 王祖望. 2001. 高原鼠兔和根田鼠的最大代谢率[J]. 动物学报, 47(6): 601-608. |
| 王祖望, 曾缙祥, 韩永才. 1979. 高原鼠兔和中华鼢鼠气体代谢的研究[J]. 动物学报, 25(1): 75-85. |
| 杨传华, 都玉蓉, 谢久祥, 等. 2012. 两种鼢鼠内脏器官形态差异及其分类学意义[J]. 兽类学报, 32(3): 259-265. |
| 叶润蓉, 曹伊凡. 1994. 高原鼠兔主要脏器重量[J]. 上海实验动物科学, 14(2): 82-84. |
| 詹纯列, 李权超, 徐本法, 等. 2001. SPF小型猪主要脏器重量脏器系数的测定[J]. 中国实验动物学杂志, 11(4): 215-217. |
| 张克勤, 陈玉梅, 刘毅, 等. 2008. 花鼠外部形态及部分内脏器官的测量和分析[J]. 四川动物, 27(6): 1104-1106, 1125. |
| 张美文, 王勇, 李波, 等. 2006. 洞庭湖区社鼠脏器重量的比较[J]. 动物学杂志, 41(1): 113-117. |
| 张世炎, 麦海. 2001. 黄毛鼠内脏器官重量和含水量的测定[J]. 动物学杂志, 36(5): 48-50. |
| 张志强, 王德华. 2009. 长爪沙鼠脏器重量和肠道长度的季节性变化[J]. 兽类学报, (3): 294-301. |
| 赵建文, 黄品杰, 付文双. 1999. 高原鼠兔解剖学特征之初探[J]. 实验动物科学与管理, 16(1): 41-44. |
| Millar JS. 1977. Adaptive features of mammalian reproduction[J]. Evolution, 32(2): 370-386. |
| Norris RW, Zhou KY, Zhou CQ, et al. 2004. The phylogenetic position of the zokors (Myospalacinae) and comments on the families of muroids (Rodentia)[J]. Molecular Phylogenetics and Evolution, 31(3): 972-978. |
| Pucek Z. 1965. Seasonal and age changes in the weight of internal organs of shrews[J]. Acta Theriologica, 10(26): 369-438. |
| Rosenmann M, Morrison P. 1974. Maximum oxygen consumption and heat loss facilitation in small homeo therms by He-O2[J]. American Journal of Physiology, 226(3): 490-495. |
| Schieck JO, Millar JS. 1985. Alimentary tract measurements as indicators of diets of small mammals[J]. Mammalia, 49(1): 93-104. |
| Schmidt-Nielsen, Knut. 1997. Animal physiology: adaptation and environment, fifth edition[M]. Cambridge: Cambridge University Press. |
| Vleck D. 1979. Energy-cost of burrowing by the pocket gopher Thomomys bottae[J]. Physiological Zoology, 52(2): 122-136. |
| Zhou C, Zhou K. 2008. The validity of different zokor species and the genus Eospalax inferred from mitochondrial gene sequences[J]. Integrative Zoology, 3(4): 290-298. |