 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 张广, 杨卫诚, 梁云, 付文泽, 张荣献, 周小利
- ZHANG Guang, YANG Weicheng, LIANG Yun, FU Wenze, ZHANG Rongxian, ZHOU Xiaoli
- 贵州白云岩喀斯特洞穴动物群落结构及其与环境因子关系研究
- Correlation Study between Cave Animal Community and the Cavern Environment in Dolomite Karst Area, Guizhou Province
- 四川动物, 2015, 34: 375-382
- Sichuan Journal of Zoology, 2015, 34: 375-382
- 10.3969/j.issn.1000-7083.2015.03.009
-
文章历史
- 收稿日期:2014-10-08
- 接受日期:2014-12-26






喀斯特洞穴是一类特殊的地下生态系统,在这类生态系统中,洞穴动物是主要组成部分,洞穴环境对洞穴动物群落结构、分布及多样性具有直接或间接的影响。在贵州喀斯特地貌分布区,有着各种复杂的洞穴,洞穴内分布着多种多样的洞穴动物。但近几年生态坏境不断恶化,对洞穴动物的分布产生了重大影响,对洞穴动物的生存造成严重威胁。
在洞穴动物及洞穴环境方面的研究,国内外均有成效,特别在国外,研究的内容广泛而深入,如区系研究( Decu & Iliffe,1983),多样性和分布研究(Wilkens et al.,2000),繁殖研究(Criffith,1991),起源和演化研究(Culver,1982),生态学和生理学研究(Biswass,1991),硫和氧对洞穴动物生活的影响研究(Sket,1996)等。在国内,某些研究领域也取得了进展,如群落研究(黎道洪等,2001),类群区系分析研究(陈德牛等,2003),与环境因子的关系研究(黎道洪,2006;张竹青等,2006;王听亚等,2007;杨卫诚,2010;黎道洪,罗蓉,2011;徐承香等,2012),对重金属的富集研究(叶子郯,黎道洪,2011;张俊等,2012)等。但国内对白云岩喀斯特地区洞穴动物分布结构与环境因子的关系研究还存在不足,特别是对贵州省施秉县白云岩地区洞穴动物的调查研究目前尚未见报道,本文通过对该地区岩孔洞和大塘下洞2个具有代表性的洞穴进行调查研究,以填补白云岩喀斯特洞穴生态系统研究的数据库,为进一步开展对洞穴动物群落结构与环境因子的关系、洞穴生态和功能的研究提供参考数据。 1 环境概况
岩孔洞位于贵州省施秉县云台山脚下居民生活区旁,居民倒入大量的生活及农业垃圾,破坏较为严重,该洞为一消水洞,洞口位于旱田旁,与旱田边缘高度差为2 m,洞口高5 m、宽3 m,海拔994 m,地理坐标108°07'28.61″E,27°08'38.92″N,洞口前有一片乱石,乱石上富含丰富的有机质,洞口处被大量地衣和落叶覆盖,洞口外分布乔木和灌木。有光带长12 m,有2个洞厅,温度为17 ℃,湿度为80%,洞底不光滑,有大量碎石,碎石上富有丰富有机质,部分区域有少量积水,洞顶不光滑,有蝙蝠栖息。弱光带长7 m,温度为15 ℃,湿度为90%,洞底积存大量湿土,洞顶不光滑。黑暗带长约20 m,温度为17 ℃,湿度为92%,黑暗带最深处为40 m2左右的水塘,具体深度不详。
大塘下洞位于贵州省施秉县云台山山脚,离居民生活区较远,该洞为一流水洞,洞口位于山谷底,地理坐标108°11'35.55″E,27°10'19.47″N,呈斜三角形,海拔1103 m,洞口高1.7 m、宽8 m,洞口周围分布大量灌木、天然细竹林及少量的蕨类植物、苔藓植物和藤本植物,整个洞穴洞底分布大量岩石和水流,黑暗带和弱光带分布有少量水塘,水塘内有水生生物分布。有光带长14 m,温度为16 ℃,湿度为90%,有光带与弱光带的过渡段有大量坍塌的岩石,弱光带长8.4 m,温度为15 ℃,湿度为92%。黑暗带长30 m,温度为16 ℃,湿度为93%,整个洞穴内分布着丰富的洞穴动物。 2 研究方法 2.1 野外调查与样品采集
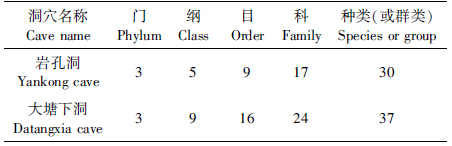
2012年8月,对施秉云台山及其附近的所有洞穴做了详细的野外考察,发现大多数洞穴都相对较浅,物种相对比较单一。而岩孔洞、大塘下洞是该区域发育比较好、洞穴动物比较丰富的洞穴,大塘下洞人为因子干扰非常少,岩孔洞受人为因素影响较大,选择这2个洞穴做该区域的代表研究,基本上能反映出该区域的洞穴物种群落结构、洞穴动物群落结构与环境的相关性以及人为破坏对洞穴动物群落结构的影响等。
野外采集过程中,用皮尺测定各光带长度、洞口宽度和高度,利用美国生产的奇遇(eTrex Venture)GPS定位仪测定海拔,利用北京亚光仪器有限责任公司生产的JWSA2-2型温湿度计测定各洞段的温湿度。光带的划分、样方确定及样品采集处理等参考黎道洪等(2001)的工作。 2.2 样品处理
对标本进行分类鉴定,由于资料缺乏、标本损坏或有些标本为幼体等原因,部分标本只能鉴定到科。环境中采集的土壤和水在室内进行分析测定,具体如下:量取250 mL水样,加入10 mL王水置温水浴中消解,消解完成后加入2.5 mL浓盐酸和5 mL 10%的硫酸定容至50 mL测定;土样烘干、研磨,过100目筛后称取0.2 g,加入10 mL的王水,置温水浴中消解,消解完成后加入2.5 mL浓盐酸和5 mL 10%的硫酸定容到50 mL测定。用AF-640原子荧光光度仪测定水样和土样中汞(Hg)和砷(As)的平均含量。 2.3 数据处理
按照Margalf的richness index公式计算物种丰富度,D=(S-1)/lnN,式中D表示物种丰富度,S表示物种数,ln为自然对数,N为各群落所有动物的总数。根据Shannon-Wiener指数公式计算群落多样性,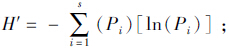 Pielou均匀性指数计算群落均匀度,J'=H'/lnS,式中H'为物种多样性指数,S为组成群落的物种或类群数,Pi为第i种物种的个体比,J'为均匀性指数。用Simpson优势度指数公式计算群落优势度,C=∑(ni/N)2,C为优势度指数,ni为第i种的个体数,N为洞内所有物种的个体数。按Whittaker的相似性指数公式分析群落的相似度,
Pielou均匀性指数计算群落均匀度,J'=H'/lnS,式中H'为物种多样性指数,S为组成群落的物种或类群数,Pi为第i种物种的个体比,J'为均匀性指数。用Simpson优势度指数公式计算群落优势度,C=∑(ni/N)2,C为优势度指数,ni为第i种的个体数,N为洞内所有物种的个体数。按Whittaker的相似性指数公式分析群落的相似度,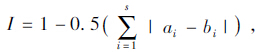 式中,I为两群落的相似性指数,s为群落中相对应的种数,ai和bi为物种或类群i的个体数分别在a和b群落中的比例。
式中,I为两群落的相似性指数,s为群落中相对应的种数,ai和bi为物种或类群i的个体数分别在a和b群落中的比例。
对岩孔洞和大塘下洞中群落多样性与部分环境因子采用Pearson相关系数进行相关性分析(张竹青等,2006;杨卫诚等,2007),并采用欧式距离公式计算其欧氏距离相关系数。通过研究洞穴土壤的单项污染指数(Pi)和综合污染指数(P)来对洞穴土壤进行重金属污染评价, 式中Pi为单项污染指数,Ci为污染物i的实测值(mg·kg-1),Si为污染物i的评价标准(mg·kg-1),P为综合污染指数,n为土壤中污染物质的种类数。单项污染指数Pi<1时,表示未污染;当1≤Pi<2时,表示轻污染;当2≤Pi<3时,表示中污染;当Pi≥3时,表示重污染。综合污染指数P<1.0时未污染,1.0~2.5轻污染,2.5~5.0中污染,5.0~10.0重污染,P>10.0严重污染(徐承香等,2010a)。
3 结果与分析
3.1 洞穴动物的种类组成和相对数量
式中Pi为单项污染指数,Ci为污染物i的实测值(mg·kg-1),Si为污染物i的评价标准(mg·kg-1),P为综合污染指数,n为土壤中污染物质的种类数。单项污染指数Pi<1时,表示未污染;当1≤Pi<2时,表示轻污染;当2≤Pi<3时,表示中污染;当Pi≥3时,表示重污染。综合污染指数P<1.0时未污染,1.0~2.5轻污染,2.5~5.0中污染,5.0~10.0重污染,P>10.0严重污染(徐承香等,2010a)。
3 结果与分析
3.1 洞穴动物的种类组成和相对数量
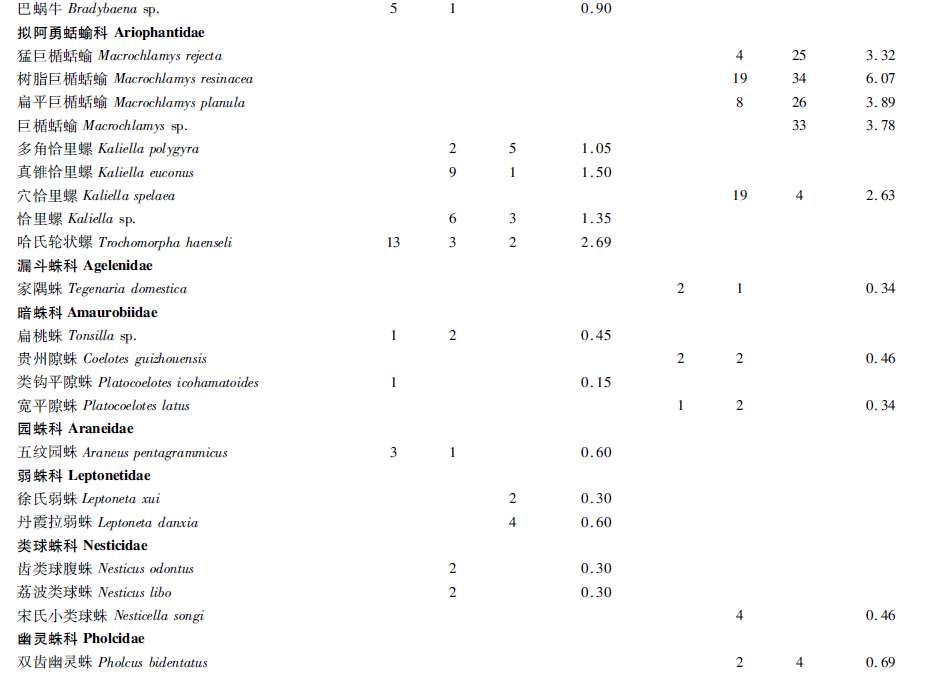
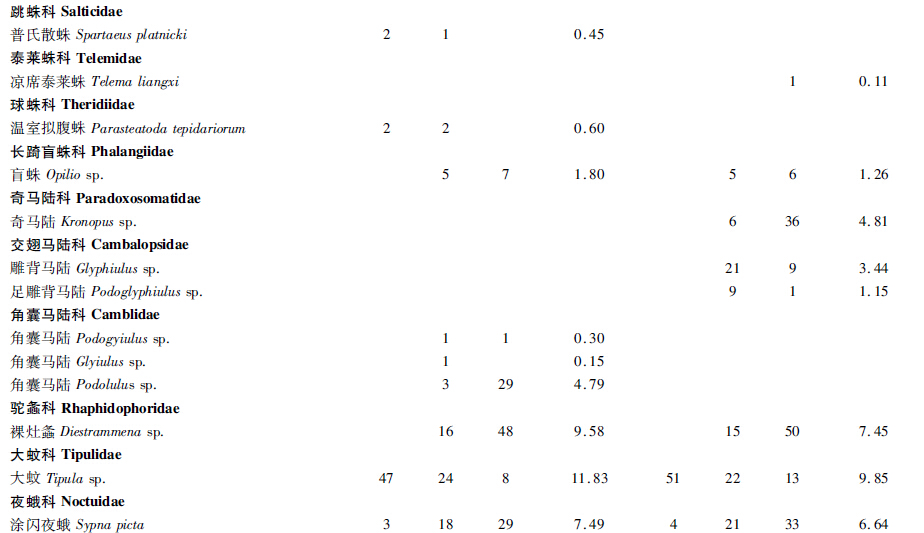
通过对2个洞穴的深入调查和标本采集,在岩孔洞共采集到标本668号,经初步鉴定隶属于3门5纲9目17科30种,大塘下洞共获标本873号,经初步鉴定隶属于3门9纲16目24科37种(表 1,表 2)。
|
|
|

|
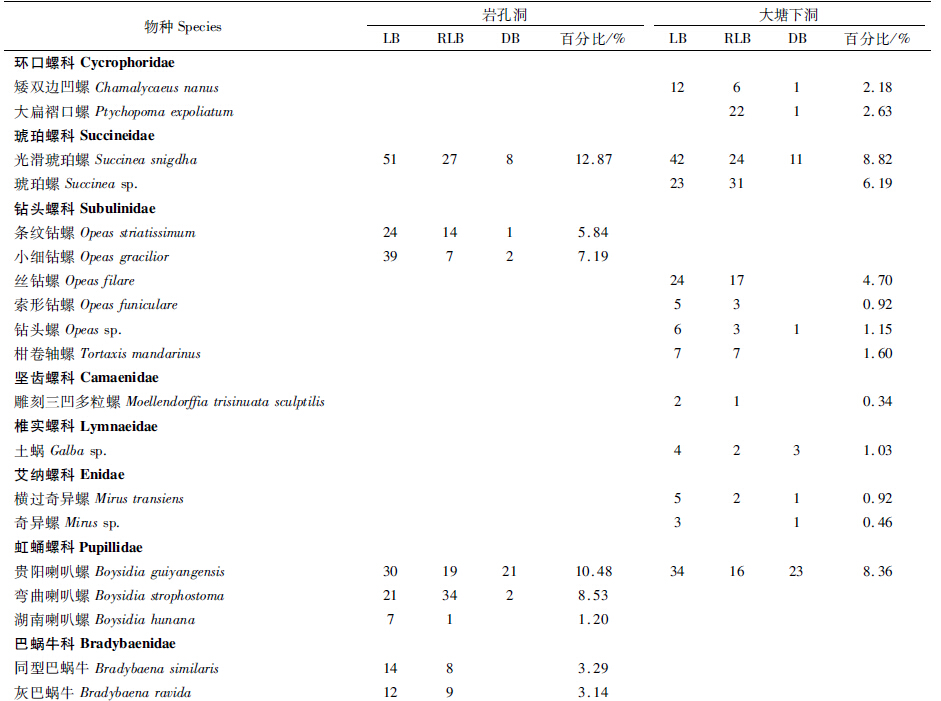
通过表 2可看出,在岩孔洞和大塘下洞内数量分布最多的物种分别是光滑琥珀螺 Succinea snigdha (12.87%)和大蚊 Tipula sp.(9.85%)。在岩孔洞中,光滑琥珀螺(12.87%)和大蚊(11.83%)为优势种,该洞优势类群较突出,而大塘下洞中,优势类群不明显。 3.2 群落组成及群落多样性分析 3.2.1 群落组成
根据2个洞穴不同光带中动物种类和数量的组成不同,及有关环境因子的差异,特别是光照强度的不同,将岩孔洞和大塘下洞的动物划分为6个动物群落,即一光带为一个动物群落。
群落A:光滑琥珀螺Succinea snigdha+大蚊Tipula sp.群落分布在岩孔洞有光带,均为优势种,分别占该光带总捕获数的18.55%和17.09%。
群落B:光滑琥珀螺Succinea snigdha+弯曲喇叭螺Boysidia strophostoma群落分布在岩孔洞弱光带,前者为优势种而后者为普通种,分别占该光带总捕获数的12.27%和15.45%。
群落C:角囊马陆Podolulus sp.+裸灶螽Diestrammena sp.+涂闪夜蛾Sypna picta群落分布在岩孔洞黑暗带,均为普通种,分别占该光带总捕获数的16.76%、27.75%和16.76%。
群落D:大蚊Tipula sp.+光滑琥珀螺Succinea snigdha群落分布在大塘下洞有光带,均为普通种,分别占该光带总捕获数的21.16%和17.43%。
群落E:光滑琥珀螺Succinea snigdha+琥珀螺Succinea sp.群落分布在大塘下洞弱光带,均为普通种,分别占该光带总捕获数的7.89%和10.20%。
群落F:奇马陆Kronopus sp.+裸灶螽Diestrammena sp.群落分布在大塘下洞黑暗带,均为普通种,分别占该光带总捕获数的10.98%和15.24%。 3.2.2 洞穴动物群落结构及群落多样性分析
多样性和均匀度指数,根据各指数计算公式计算出6个群落的多样性指数(表 3)。
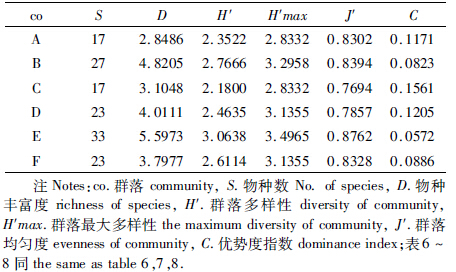
从表 3可得出,丰富度指数最高的是群落E(5.5973),分布在大塘下洞弱光带,丰富度指数体现群落总体的物种数和个体数,物种数越多,丰富度指数就越大,而个体数越多,丰富度指数反而变小,此结果与前人研究相同(杨卫诚等,2013)。均匀度指数由高到低的顺序是群落E(0.8762)>B(0.8394)>F(0.8328)>A(0.8302)>D(0.7857)>C(0.7694)。群落多样性指数最高的是群落E(3.0638),分布在大塘下洞弱光带,该指数体现群落物种数以及物种分布的均匀程度,物种数越多,均匀度越高,群落多样性指数也就越高。物种优势度从高到低的顺序是群落C(0.1561)>D(0.1205)>A(0.1171)>F(0.0886)>B(0.0823)>E(0.0572)。在2个洞穴中,除物种优势度外,弱光带物种和群落多样性均大于有光带和黑暗带。在6个群落中,群落E的物种和群落多样性最高,均匀度也最高,而物种优势度最小,分布较均匀。导致该结果的原因可能是,在这个洞穴中有光带和弱光带相对较长,又因为是水洞,在雨水季节大量洞外腐殖质被带入洞内,使这一小环境中的食物资源相对较丰富,较适宜更多半洞动物的生存。而有光带与外界接触较广,生存于此的物种易受到外界生物的影响,如鸟类捕食等,长期以来,生存于弱光带的群落E就有更优越的生态环境。
不同群落间相似性指数见表 4。通过表 4可得出,同一洞穴相邻2个群落间相似性指数相对较高,如群落A-B(0.6209)>A-C(0.253)。分布在不同洞穴或同一洞穴隔离较远的2个群落之间相似性指[JP3]数相对较低,如群落A-F(0.1542),群落D-F(0.1854)。群落间的相似性在很大程度上受环境因子影响,2个群落生存的生态环境越接近或越相似,其构成可能会越接近。

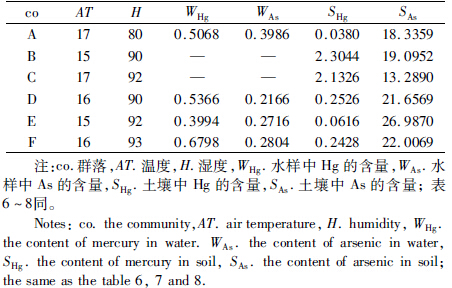
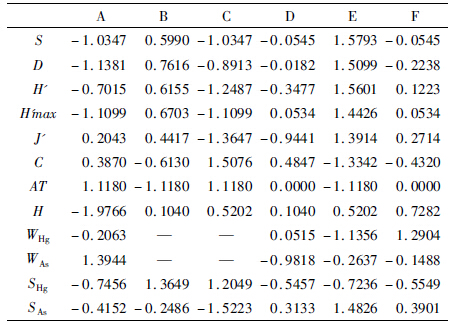
在前人研究中,湿度和温度对洞穴动物分布影响较大(徐承香等,2012)。重金属Hg的毒性较其他重金属强,元素As存在于多种农药中,且毒性较强(徐争启等,2008)。在调查研究过程中,分别对2个洞穴每个光带的温度、湿度、土壤及水中Hg和As平均含量进行测定和检测,Hg和As含量单位为mg·kg-1,测定值见表 5。利用SPSS 17.0统计分析软件分析群落多样性及其与环境因子的相关性。由于多样性与环境因子单位不统一,因此对表 3和表 5中的数据进行标准差标准化处理,消除量纲影响后所得值见表 6。
|
|
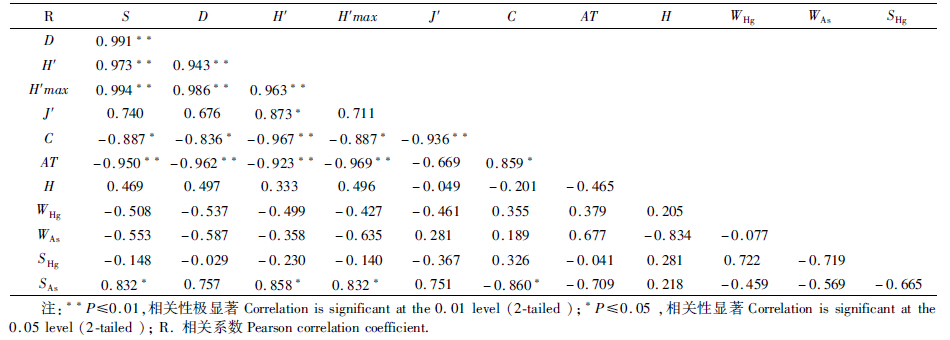
利用SPSS 17.0统计分析软件对表 6中的数据进行Pearson相关分析,得到表 7中各种相关系数矩阵。
|
从表 7可知,在物种或群落多样性指数的相关性分析中,除J'与S、D、H'max相关性不显著外,其余各群落多样性指数之间相关性较显著。从环境因子与物种或群落多样性相关性分析来看,AT 与S、D、H'、H'max呈极显著负相关,相关系数分别为-0.950、-0.962、-0.923、-0.969(P≤0.01),AT 与C呈显著正相关,相关系数为0.859(P≤0.05)。SAs与S、H'、H'max呈显著正相关,相关系数分别为0.832、0.858、0.832(P≤0.05),SAs与C呈显著负相关,相关系数为-0.860(P≤0.05)。其余的相关性均不显著。分析表明,在2个洞中群落多样性指数间不是孤立的,而是相互关联、相互影响的。洞穴环境对洞穴动物的分布存在一定程度的影响,如温度在很大程度上影响着洞穴动物分布,重金属As对洞穴动物的分布也产生一定程度的影响。 3.4 洞穴动物群落多样性与环境因子的欧氏距离分析
利用SPSS 17.0统计分析软件对岩孔洞和大塘下洞洞穴动物群落多样性、部分环境因子进行欧式距离分析,得到动物群落的各种相关系数距离分析矩阵(表 8)。

|
从表 8可知,在岩孔洞和大塘下洞中,除湿度与其他各系数间欧氏距离较大外,其余均较小,SHg与C欧氏距离最小,仅为0.22。表明所研究的洞穴动物群落之间、环境因子之间、洞穴动物与环境因子之间在很大程度上相互关联,相互制约,相互影响。
从表 7和表 8的分析可以得出,群落的相关分析和距离分析存在一定的差异,如SHg与C进行Pearson相关分析时,相关性不显著,而欧氏距离分析时,欧氏距离只有0.22,两者相关性较显著。又如环境因子在进行Pearson相关分析时,相关性均不显著,但在欧氏距离分析时,除湿度(H)与其他因子欧氏距离较大外,其余因子间欧氏距离均较小,即相关性较显著。出现这种结果的原因主要是分析方法的侧重点不同,Pearson相关分析是分析2个变量间的线性关系程度,而欧氏距离分析则是分析2个变量或2个观测量间的相似或不相似程度。使用2种方法进行综合分析,能够更客观地反映各参数的实效性和有用性。分析结果表明,洞穴中的群落间不是孤立存在的,都存在着直接或间接的联系,特别是个体与个体之间、物种与物种之间、群落与群落之间、群落与环境之间都通过食物网联系在一起,在很大程度上相互制约、相互影响。 3.5 土壤重金属Hg和As的污染评价
通过计算,岩孔洞和大塘下洞土壤中重金属Hg和As的污染指数见表 9。

|
通过表 9可知,在岩孔洞内,Hg的单项污染指数平均值显示为重污染,污染指数为9.944,这说明Hg对岩孔洞污染较严重;从综合污染指数看,除有光带外,均显示为重污染。有光带土壤中Hg显示为未污染,可能是受雨季水流作用稀释等引起的。在弱光带和黑暗带,随流水积累作用、人为干扰等多种原因,重金属含量升高。总体看,岩孔洞内Hg和As污染较严重,特别是Hg污染非常严重。在大塘下洞内,单项污染指数和综合污染指数在每个光带内均显示为轻污染,这表明大塘下洞受到Hg和As的污染,但污染不严重。在调查研究的2个洞内,土壤均受到Hg和As的污染,从表 7和表 8分析可知,Pearson相关分析时,SAs与S、H'、H'max均呈显著正相关,与C呈显著负相关;欧氏距离分析时,SHg与C欧氏距离仅为0.22,与其他群落多样性指数欧氏距离也较小。这表明洞穴土壤重金属与洞穴动物分布存在显著相关性,即在一定程度上严重影响洞穴动物分布,对洞穴动物生存产生严重影响。而洞穴土壤受到重金属污染的原因主要有地表重金属伴随地表水通过岩层渗入洞穴土壤,洞穴土壤母质中的重金属,洞口流水、动物活动和人为活动等带入。 4 小结
在不同的洞穴中,动物群落均存在着一定的差异性,但在很大程度上都受环境因子及人类活动的综合影响。本文中研究的2个洞穴,在欧氏距离和Pearson相关性分析中,都能表明环境因子对洞穴动物群落结构的综合影响。另外,在Pearson相关性分析中,温度对群落多样性均有显著相关关系,这与前人研究的有所不同(黎道洪,2006;徐承香等,2010b;杨卫诚等,2013),这可能是前人研究的洞穴基本上都为典型的石灰岩发育,与本文研究的白云岩洞穴存在着一定的地质结构差异,这有待于后续研究。另外,土壤重金属污染在一定程度上严重影响着洞穴动物的分布结构及群落多样性,如岩孔洞的污染等级相对较高,动物群落多样性指数就显著低于大塘下洞。所以,需要更进一步地摸清白云岩地区洞穴动物的群落结构,全面提升对贵州施秉白云岩喀斯特洞穴生态系统中动物群落以及其生态功能研究,更好地保护白云岩喀斯特这一特殊洞穴动物资源。
| 陈德牛, 黎道洪, 罗太昌. 2003. 贵州洞穴淡水、陆生贝类区系及新种记述[J]. 动物分类学报, 28(4): 614-620. |
| 黎道洪, 罗蓉, 陈浒. 2001. 贵州大洞口内若干动物群落研究[J]. 生态学报, 21(1): 126-130. |
| 黎道洪, 罗蓉. 2011. 贵州龙洞内动物群落结构和分布与部分环境因子的关系研究[J]. 中国岩溶, 20(4): 315-320. |
| 黎道洪. 2006. 贵州水拔洞内动物群落结构与部分环境因子的相关性研究[J].中国岩溶, 25(3): 256-262. |
| 黎道洪. 2007. 贵州波多洞和甲良洞内部分环境因子与动物群落结构的相关性[J]. 生态学报, 27(6): 2167-2176. |
| 王听亚, 李廷勇, 胡蓉, 等. 2007. 重庆芙蓉洞滴水地球化学初探[J]. 西南大学学报: 自然科学版, 29(2): 122-126. |
| 徐承香, 李子忠, 黎道洪. 2012. 喀斯特洞穴动物与部分生态环境因子的关系[J]. 湖北农业科学, 51(6): 1081-1084. |
| 徐承香, 黎道洪, 梅杰. 2010a. 贵州郑家小洞内动物重金属含量及与土壤和水体的研究[J]. 中国岩溶, 29(1): 48-54. |
| 徐承香, 黎道洪, 梅杰. 2010b. 贵州大水洞和马狼洞内动物群落结构与部分环境因子的相关性研究[J]. 贵州师范大学学报: 自然科学版, 28(2): 1-9. |
| 徐争启, 倪师军, 庹先国, 等. 2008. 潜在生态危害指数法评价中重金属毒性系数计算[J]. 环境科学与技术, 31(2): 112-115. |
| 杨卫诚, 黎道洪, 徐承香. 2013. 广西大岩洞和岩头2号洞洞穴动物群落结构及其与环境因子关系分析[J]. 四川动物, 32(3): 442-448. |
| 杨卫诚, 黎道洪. 2010. 广西大岩洞洞穴动物体与环境中重金属含量关系[J]. 生态学杂志, 29(8): 1554-1558. |
| 杨卫诚, 黎道洪, 苏晓梅. 2007. 贵州紫云洞和樱桃洞洞内动物的群落结构与洞穴环境关系初步研究[J]. 四川动物, 26(4): 738-745. |
| 叶子郯, 黎道洪. 2011. 贵州屯上洞洞穴动物对5种重金属富集的研究[J]. 四川动物, 30(3): 372-376. |
| 张俊, 黎道洪, 杜典松. 2012. 贵州龙井洞和白龙洞裸灶螽对重金属的富集[J]. 广西师范大学学报: 自然科学版, 30(4): 104-109. |
| 张竹青, 黎道洪, 罗卫东. 2006. 贵州姜家洞不同光带内动物群落结构和分布与洞穴环境的关系[J]. 贵州师范大学学报: 自然科学版, 24(3): 1-6. |
| Biswass J. 1991. Metaboliciency and regulation of body weight: a comparison life in hypogen and ecosystem[J]. Int J Speleol, 20(1): 15-22. |
| Criffith DM. 1991. The effects of substrate moisture on survival of adult cave beetles (Neaphaenops tellkampfi) and cave cricket eggs (Hadenoecus subterraneus)in a sandy deep cave sit[J]. NSS Bull, 53(2): 98-103. |
| Culver DC. 1982. Cave life: evolution and ecology[M]. Cambridge: Harvard University Press: 189. |
| Decu VG, Iliffe TM. 1983. A review of theter restrial cavemieolous fauna of Romania[J]. NSS Bulletin, 45: 86-97. |
| Sket B. 1996. The ecology of anchihaline caves[J]. Trends in Ecology & Evolution, 11(5): 221-225. |
| Wilkens H, Culver DC, Humphreys WF. 2000. Ecosystems of the world 30: subterranean ecosystems[M]. Amsterdam: Elsevier. |