 2015, Vol. 34(2)
2015, Vol. 34(2)扩展功能
文章信息
- 李佳, 李言阔, 缪泸君, 谢光勇, 袁芳凯
- LI Jia, LI Yankuo, MIAO Lujun, XIE Guangyong, YUAN Fangkai
- 桃红岭国家级自然保护区梅花鹿和野猪秋季生境选择差异
- A Comparison of Autumn Habitat Selection of Cervus nippon and Sus scrofa in Taohongling National Nature Reserve, China
- 四川动物, 2015, 34(2): 300-305
- Sichuan Journal of Zoology, 2015, 34(2): 300-305
- 10.3969/j.issn.1000-7083.2015.02.023
-
文章历史
- 收稿日期:2013-10-30
- 接受日期:2014-10-17






生境选择是反映动物对资源(如食物、空间、水源等)利用差异的一个重要因素,不同动物有自己独特的资源利用方式(Henley et al.,2007)。研究表明,在资源有限的生境中,生境选择的差异是同域分布的野生动物生态位分歧的最主要形式(Wei et al.,2000),也是同域物种共存的基础(Schoener,1974)。因此,通过了解生境选择的差异,解释同域物种之间的资源分配方式已经成为群落生态学领域重要的研究内容(骆颖等,2010)。近年来,同域物种生境选择的差异得到了比较广泛的研究,尤其是偶蹄类动物,包括岩羚羊Rupicapra rupicapra和盘羊Ovis musimon(Gaelle et al.,2012)、野驴Equus hemionus和瞪羚Gazella dorcas(Henley et al.,2007)、塔尔羊Hemitragus jemlahicus和岩羚羊(Forsyth,2000)、岩羊Pseudois nayaur和马鹿Cervus elaphus(骆颖等,2010)、矮岩羊Pseudois schaeferi和斑羚Naemorhedus goral(申定健等,2009)、小麂Muntiacus reevesi和野猪Sus scrofa(鲁庆彬等,2007)等物种。
梅花鹿Cervus nippon属偶蹄目Artiodactyla鹿科Cervidae鹿属Cervus,是东亚季风区特有的鹿类,是国家Ⅰ级重点保护野生动物,曾有6个亚种广泛分布于我国东北、华北、华中、华南、西南和青藏区的东部(郭延蜀,郑惠珍,2000)。野猪属偶蹄目猪科Suidae野猪属Sus,中国仅存1个野猪种,有7个亚种,几乎遍布全国(张劲硕,2007)。在江西桃红岭梅花鹿国家级自然保护区,两者同域共存(蒋志刚,2009)。梅花鹿为草食性动物,常年以各种植物为食,采食种类很多(姚振生,2010);野猪为杂食性(Fournier et al.,1995),喜食一些含淀粉、糖和脂肪较多的食物,因此,它们具有相似的食物需求,长期共同进化的结果可能使它们在生境选择方面存在一定的差异。开展梅花鹿和野猪生境选择的研究,既可以为促进该地物种多样性保护提供参考,亦有助于了解物种间同域共存的机制。秋季是梅花鹿发情交配的季节(江西桃红岭梅花鹿保护区管理处,2000),此时是梅花鹿重要的生理时期,研究梅花鹿和野猪在这个时期的生境选择及其差异,对了解野猪对梅花鹿的繁殖和生存策略的影响具有重要意义。为揭示梅花鹿和野猪的生境利用方式,作者于2012年10—12月对保护区的梅花鹿和野猪进行了生境选择研究,旨在为维护梅花鹿和野猪这两个种群的生态平衡提供科学依据。 1 研究区域自然概况
江西桃红岭梅花鹿国家级自然保护区(29°42'~29°53'N,116°32'~116°43'E)位于长江中下游南岸,江西省最北沿彭泽县的中部。总面积12 500 hm2,其中核心区2670 hm2,实验区1830 hm2,缓冲区8000 hm2。地貌为平缓起伏的低山丘陵,海拔100~500 m,最高峰猫鹰窝海拔536.6 m。气候属亚热带季风类型,日照充足,雨量充沛,年平均降水量1300 mm,年平均气温为15.1 ℃,年均日照时数2043.6 h。保护区内植被根据群落外貌和结构特征可划分为针叶林、常绿阔叶林、针阔混交林、落叶阔叶林、灌丛、竹林、芭茅丛、草丛(李佳等,2014)(图 1)。主要乔木树种有乌楣栲Castanopsis jucunda、马尾松Pinus massoniana、杉树Cunninghamia lanceolata、石栎Lithocarpus glaber、化香Platycarya strobilacea、朴树Celtis sinensis等。主要灌木和草本植物有美丽胡枝子Lespedeza formosa、盐肤木Rhus chinensis、映山红Rhododendron simsii、白茅Imperata cylindrica、葛藤Pueraria lobata、悬钩子Rubus palmatus、五节芒Miscanthus floridulus等(江西桃红岭梅花鹿保护区管理处,2000)。

|
| 图 1 江西桃红岭梅花鹿国家级自然保护区植被类型分布图 Fig. 1 The vegetation in Taohongling Sika Deer National Nature Reserve,Jiangxi province |
梅花鹿和野猪均是森林林栖动物,野外很难直接观察,我们主要利用样方法记录样方内梅花鹿和野猪的粪便、食痕、蹭迹、足迹、卧迹(窝)等新鲜活动的痕迹,判定它们对生境的利用情况。 2.1 样方设置
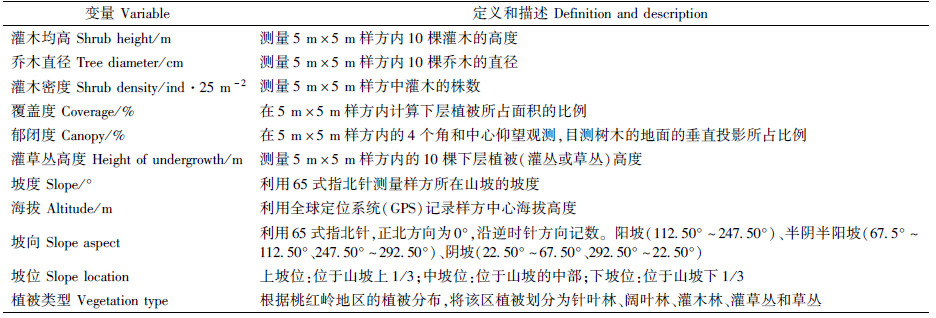
本研究于2012年10—12月采用样线法和样方法对保护区内梅花鹿和野猪栖息地进行了野外调查。利用1∶[KG-2mm]50 000地形图,随机布设样线,所设置样带覆盖了保护区内主要生境类型,在样线上按水平距离每隔100 m机械布点的方法设置5 m×5 m的样方,共布设16条样线,样线间距>1000 m,长度>1500 m,在样线上共设置样方312个,其中梅花鹿利用样方89个,野猪利用样方112个,对照样方111个。在样方内发现梅花鹿或野猪的新鲜活动痕迹时,记录痕迹发现点的灌木均高、乔木直径、灌木密度、覆盖度、郁闭度、灌草丛高度、坡度、海拔、坡向、坡位、植被类型共11类生态因子,生态因子的测定方法见表 1。
用Chi-square检验分析梅花鹿和野猪在植被类型、坡向和坡位等3类非数值型生态因子的选择上是否存在差异。通过Kolmogorov-Smironov Test检验灌木均高、乔木直径、灌木密度、覆盖度、郁闭度、灌草丛高度、坡度、海拔等8类数值型生态因子的数据是否呈正态分布,如果不符合正态分布,则采用Kruskal-Wallis H Test进行检验。对于各样方组差异存在统计学意义的变量,则通过Mann-Whitey U Test比较组间生境变量上的差异,以获取梅花鹿和野猪的生境利用模式。对于8个数值型生态因子进行判别分析,由于数据不符合正态分布,因此先对数据利用Johnson分布体系进行标准化,然后采用逐步判别分析对梅花鹿和野猪的生态因子进行分析,以揭示影响它们生境选择差异的关键因子。所有数据均利用SPSS 17.0进行分析,显著性水平α=0.05。 3 结果 3.1 梅花鹿和野猪在生态因子选择上的差异
Chi-square检验表明,秋季梅花鹿和野猪对植被类型、坡向和坡位的选择差异均有高度的统计学意义(植被类型:χ2=90.64,df=4,P<0.001;坡向: χ2=16.55,df=2,P=0.004;坡位:χ2=63.32,df=2,P<0.001)。梅花鹿选择草丛和灌丛、坡向为阳坡、坡位为上坡位的生境;而野猪则偏爱灌木林生境,随机选择植被类型、坡向和坡位。
Kruskal-Wallis H Test检验结果表明,梅花鹿利用样方、野猪利用样方和对照样方在海拔、灌草丛高度、灌木密度、坡度和灌木均高等5个变量上的差异都有统计学意义,而对乔木直径、郁闭度、覆盖度等变量的差异则无统计学意义。结合变量在各组中的平均值可以看出,梅花鹿频繁出现在高海拔(365.5 m±141.7 m)、坡度平缓(17.80°±12.68°)、灌丛或草丛高度较矮(0.81 m±0.36 m)、灌木密度较小(7.56棵±4.55棵)、高度较高(4.70 m±1.68 m)的生境;而野猪对所有的变量表现为随机选择,差异无统计学意义(表 2)。

在梅花鹿-野猪组判别函数中,坡度、海拔和灌木密度3个变量的判别函数系数的绝对值明显大于其他变量(表 3),表明主要由这3个变量导致了梅花鹿与野猪之间的生境分离(Wilk's λ=0.801,χ2=24.89,df=3,P<0.001),区分梅花鹿与野猪样方的判别函数方程:F=0.653×坡度-0.546×海拔+0.840×灌木密度+0.144,正确判别率达82.4%。类似的坡度、灌木密度和乔木直径是梅花鹿利用样方与对照样方生境分离的主要原因(Wilk's λ=0.765,χ2=28.22,df=3,P<0.001),区分梅花鹿利用样方与对照样方的判别函数方程:F=0.587×坡度+0.639×乔木直径+0.705×灌木密度+0.000,正确判别率达70.1%。而在野猪利用样方与对照样方之间并无显著判别函数系数的变量(Wilk's λ=0.972,χ2=3.83,df=8,P=0.872)。

然而,仅3个变量,即坡度、乔木直径和灌木密度进入了最后的逐步判别函数方程中(表 4)。这种结果的出现可能与选择变量进入判别方程的当量值标准有关(>3.84进入方程,<2.71则从方程剔除)。由于逐步判别函数分析只保留具有相对更大贡献的变量,因此这3个变量相对于其他变量在判别各组样方中具有更大的贡献能力。共建立2个判别函数,由这3个变量区分秋季梅花鹿利用样方、野猪利用样方和对照样方的正确率达到72.2%。以2个判别函数建立的判别分类图形象地揭示了梅花鹿利用样方、野猪利用样方和对照样方存在一定程度的重叠和分离(图 2),其中,梅花鹿利用样方明显偏离了对照样方,并且与野猪的重叠较少,而野猪利用样方则与对照样方重叠较多。


|
| 图 2 梅花鹿利用样方、野猪利用样方和对照样方的散点图 Fig. 2 Scatter plot for sika deer quadrats,wild boar quadrats and control quadrats |
某一特定的动物物种总是出现在其喜好的生境中。马建章等(2004)认为动物的生境可以指某一种动物生存所需的空间,亦可以指许多种动物生活的环境。动物对生境的选择往往受到各种环境因素的综合作用,但许多研究表明动物种间或种内竞争因素要比生境本身更加影响动物对生境的选择(Crowell & Pimm,1976;Hairston,1981)。梅花鹿和野猪都集中在秋季发情、交配,为寻找伴侣,其活动范围比其他季节更大(郭延蜀等,1991;王文等,2007),种内和种间竞争亦趋激烈。因此,调查梅花鹿与野猪这2个物种在发情季节对生境的利用情况,对探讨其同域共存的机制具有一定意义。
竞争排斥原理认为:生态位上相同的2个物种,不能在同一区域内长期共存,如果要在同一区域内共存,它们之间必然在栖息地、食性、活动时间或其他特征上发生生态分化(孙儒泳,2001)。在桃红岭梅花鹿国家级保护区,梅花鹿和野猪均在晨昏采食,晚上卧息,喜食多汁幼嫩的植物(杨晶等,1990;高中信,王文,2007;姚振生等,2010),表明这2个物种在食性和活动时间上重叠,因此我们预测生境选择的差异是其分离的主要原因。本文所设定的11种生态因子中,野猪和梅花鹿秋季在植被类型、灌草丛高度、灌木密度、坡度、灌木均高、坡向、坡位和海拔8个生态因子上的选择差异均具有统计学意义。逐步判别分析的结果也同样显示梅花鹿和野猪在生境选择上几乎完全分离,从而证明通过生境分离来实现对资源的平衡分配是梅花鹿和野猪的选择策略之一,这与预测吻合。这种生境选择的差异有利于降低梅花鹿与野猪之间的相互干扰和资源竞争,保证了它们各自的需求,以便两者更好地共存。
近年来,桃红岭地区野猪种群数量剧增,除与其极强的繁殖能力外,还与其对生境的适应能力有关。从研究结果可以看出,桃红岭地区野猪对生境的选择较为随机,说明其对各种生境有较强的适应能力,整个地区几乎都能看到野猪活动的痕迹,活动范围较大。与此形成鲜明对比的是梅花鹿的活动范围,由于其生性胆怯、易惊,对生境的要求极其严格,主要选择海拔300 m以上的空间活动。因此,我们认为野猪对生境的适应能力远远强于梅花鹿,野猪与梅花鹿争夺栖息地、食物和水源,压缩梅花鹿的生存空间,这与付义强等(2006)、吴问国等(2008)的研究结果一致,认为不断扩大的野猪种群对梅花鹿种群的发展产生了不利影响,是造成梅花鹿种群增长较慢的原因之一。
在不同的地区,野猪的生态作用是不同的。研究者从不同的研究角度揭示了野猪这一大型杂食类哺乳动物具有的不同生态作用(滚双宝,2004)。因此,野猪对生态环境的作用方式在很大程度上取决于当地的资源特点。桃红岭保护区的主要保护对象为梅花鹿,因此,在评定野猪的保护价值和生态作用时,必须充分考虑野猪对梅花鹿生存的影响,建议在野猪管理工作中,管理部门需对其进行持续的监控,避免野猪种群泛滥成灾,最大限度减少其与梅花鹿发生冲突。
致谢:野外调查过程中得到了江西桃红岭国家级自然保护区生态监测站工作人员的大力支持,在此表示衷心的感谢。| 付义强, 胡锦矗, 郭延蜀, 等. 2006. 桃红岭自然保护区梅花鹿对春季栖息地的利用[J]. 动物学杂志, 41(4): 60-63. |
| 高中信, 王文. 2007. 野猪[J]. 生物学通报, 42(1): 6-7. |
| 滚双宝. 2004. 野猪种群特性及其生态作用的研究进展[J]. 特产研究, 1: 52-56. |
| 郭延蜀, 胡锦矗, 罗代华, 等. 1991. 四川梅花鹿的社群行为研究[J]. 兽类学报, 11(3): 165-170. |
| 郭延蜀, 郑惠珍. 2000. 中国梅花鹿地史分布、种和亚种的划分及演化历史[J]. 兽类学报, 20(3): 168-179. |
| 江西桃红岭梅花鹿保护区管理处. 2000. 江西桃红岭梅花鹿保护区[M]. 北京: 中国林业出版社. |
| 蒋志刚. 2009. 江西桃红岭国家自然保护区生物多样性[M]. 北京: 清华大学出版社. |
| 李佳, 李言阔, 缪泸君, 等. 2014. 江西桃红岭国家级自然保护区梅花鹿适宜性评价[J]. 生态学报, 34(5): 1274-1283. |
| 鲁庆彬, 于江傲, 高欣, 等. 2007. 冬季清凉峰山区小麂和野猪的生境选择及差异[J]. 兽类学报, 27(1): 45-52. |
| 骆颖, 张明明, 刘振生, 等. 2010. 贺兰山同域分布岩羊和马鹿发情季节的生境选择差异[J]. 生态学报, 30(14): 3744-3751. |
| 马建章, 邹红菲, 贾竞波. 2004. 野生动物管理学[M]. 哈尔滨: 东北林业大学出版社. |
| 申定健, 郑合勋, 王淯, 等. 2009. 四川省巴塘县矮岩羊与斑羚冬季生境比较[J]. 生态学报, 29(5): 2321-2330. |
| 孙儒泳. 2001. 动物生态学原理(第三版)[M]. 北京: 北京师范大学出版社. |
| 王文, 张静, 马建章. 2007. 小兴安岭南坡野猪家域分析[J]. 兽类学报, 27(3): 257-262. |
| 吴问国, 朱文, 高依敏, 等. 2008. 江西桃红岭野生梅花鹿保护现状及管理对策[J]. 野生动物, 27(3): 457-459. |
| 杨晶, 丁铁明, 胡平喜, 等. 1990. 梅花鹿南方亚种生态学研究初报[J]. 野生动物, 11(3): 17-19. |
| 姚振生, 徐向荣, 徐攀. 2010. 江西桃红岭梅花鹿华南亚种食源植物分析[R]. 全国第9届天然药物资源学术研讨会: 236-331. |
| 张劲硕. 2007. 世界猪的种、分布和现状[J]. 动物学杂志, 42(2): 7-19. |
| Crowell KL, Pimm SL. 1976. Competition and niche shifts of mice introduced onto small islands[J]. Oikos, 27(2): 251-258. |
| Forsyth DM. 2000. Habitat selection and coexistence of the Alpine chamois (Rupicapra rupicapra) and Himalayan tahr (Hemitragus jemlahicus) in the eastern southern Alps, New Zealand[J]. Journal of Zoology, 252(2): 215-225. |
| Fournier CC, Maillard D, Fournier P. 1995. Diet of the wild boar (Sus scrofa L.) inhabiting the Montpellier garrigue[J]. Ibex, 3: 174-179. |
| Gaelle D, Clement C, Anne L, et al. 2012. Spatial distribution and habitat selection in coexisting species of mountain ungulates[J]. Ecography, 35(1): 44-53. |
| Hariston NG. 1981. An experimental test of a guild: salamander competition[J]. Ecology, 52(1): 65-72. |
| Henley SR, Ward D, Schmidt I. 2007. Habitat selection by two desert-adapted ungulates[J]. Journal of arid environments, 70(1): 39-48. |
| Schoener TW. 1974. The compression hypothesis and temporal resource partitioning[J]. Proceedings of the National Academy of Sciences of the United States of America, 71(10): 4169-4172. |
| Wei FW, Feng Z, Wang Z, et al. 2000. Habitat use and separation between the giant and the red panda[J]. Journal of Mammal, 80(2): 448-455. |