 2015, Vol. 34(2)
2015, Vol. 34(2)扩展功能
文章信息
- 金星星, 张耀光, 陈幕飞, 金丽*
- JIN Xingxing, ZHANG Yaoguang, CHEN Mufei, JIN Li*
- 4种两栖爬行动物血细胞的显微结构及细胞化学特征观察
- Observations of Microscopic Structures and Cytochemical Characteristics of Blood Cells in Four Species of Amphibians and Reptiles
- 四川动物, 2015, 34(2): 223-228
- Sichuan Journal of Zoology, 2015, 34(2): 223-228
- 10.3969/j.issn.1000-7083.2015.02.011
-
文章历史
- 收稿日期:2014-05-31
- 接受日期:2014-10-17






血液是动物体内一种极其重要的组织,通过血液学指标可初步诊断疾病和健康状况,为动物生理病理毒理学研究提供依据。目前已有很多关于两栖类和爬行类外周血细胞显微结构的研究文献(郭宪光等,2002;黄斌等,2002;胡知渊等,2005;李桂芬等,2009;周庆萍等,2010),但关于两栖类和爬行类外周血细胞的细胞化学特征方面的研究极少(王石泉,1995)。因此我们对两栖动物中华蟾蜍Bufo gargarizans和牛蛙Rana catesbeiana,爬行动物铜蜓蜥Sphenomorphus indicus和青海沙蜥Phrynocephalus vlangalii外周血细胞的细胞化学染色特征进行了观察与比较,旨在充实其生物学资料,丰富动物血液生理、比较细胞学等方面的资料。 1 材料与方法 1.1 材料
实验用中华蟾蜍、牛蛙、铜蜓蜥和青海沙蜥,各5只。牛蛙和青海沙蜥购自重庆市北碚某市场,中华蟾蜍和铜蜓蜥采自西南大学校园,均为健康成体。 1.2 方法 1.2.1 血细胞涂片制备
采血时,打开围心腔,用已抗凝的注射器从心脏取血后立即涂片。部分涂片经甲醇固定后做常规Wright-Giemsa染色,观察显微结构;部分涂片做细胞化学染色,观察细胞化学特征。 1.2.2 细胞化学染色
苏丹黑B(SBB)染色显示脂类物质,偶氮偶联法显示酸性磷酸酶(ACP)和碱性磷酸酶(AKP)(龚志锦等,1994)。双氨基联苯胺法显示过氧化物酶(POX),酸性-α-醋酸萘酚染色显示酸性-α-醋酸萘酯酶(ANAE)(刘志洁等,2002)。后4种染色的阴性对照均采用不含底物的孵育液处理。每个个体每种染色处理6张涂片。1.2.3 染色结果评价参照Tavares-Dias(2006)的方法将阳性程度划分为3个等级:阳性区域占整个细胞质面积3/4以上划分为强阳性(+++);占1/2为阳性(++);小于1/2为弱阳性(+)。没有阳性区域为阴性(-);同一类染色的所有涂片上没有观察到此类细胞,为缺乏此类血细胞(CA)。染色越深代表阳性反应越强。1.3 测量和数据处理用Nikon 80i研究型显微镜观察各类血细胞形态和细胞化学染色特征并拍照。用IPP图像处理软件进行细胞大小测量。每个个体每种细胞随机选取≥50个测量大小。结果用平均值±标准差(Mean±SD)表示,并对同一种细胞在不同动物的大小用SPSS 17.0中Duncan,s多重比较方法进行统计分析(α=0.05)。 2 结果 2.1 显微结构
4种动物外周血中均可观察到红细胞、单核细胞、淋巴细胞、血栓细胞、嗜中性粒细胞和嗜碱性粒细胞,中华蟾蜍和牛蛙的外周血中还观察到嗜酸性粒细胞。这4种动物外周血细胞形态相似,但大小存在差异(表 1)。

|
红细胞长椭圆形,表面光滑,核中位。青海沙蜥和中华蟾蜍的外周血中均可见无核红细胞(图版Ⅰ:1)。血栓细胞圆形、椭圆形或长棒状,胞质较少,在核外围成极薄的一圈(图版Ⅰ:2)。大淋巴细胞(图版Ⅰ:3)圆形或椭圆形,表面有许多突起;核大近圆形,胞质少。小淋巴细胞(图版Ⅰ:4)形态与大淋巴细胞相似,胞质极少。单核细胞卵圆形,核呈椭圆形、肾形、马蹄形等,胞质丰富,常可见有伪足状突起。内有少量嗜天青颗粒及大小不等的空泡。青海沙蜥外周血中还可见巨噬细胞(图版Ⅰ:5)。嗜中性粒细胞卵圆形或椭圆形,核圆形、椭圆形、肾形、分叶形(图版Ⅰ:6),胞质几乎不着色,内有淡蓝色丝状或颗粒状结构。嗜碱性粒细胞数目稀少,细胞圆形或椭圆形,胞核椭圆形,分叶不明显,胞浆中充满深蓝黑色圆形或菱形颗粒(图版Ⅰ:7)。嗜酸性粒细胞圆形或椭圆形,核常椭圆形或二分叶,偶见2个以上核;胞浆中充满均一整齐的圆形或椭圆形桔红色嗜酸性颗粒(图版Ⅰ:8),只在中华蟾蜍和牛蛙的外周血中观察到嗜酸性粒细胞。

|
| 图版Ⅰ3种两栖爬行动物外周血细胞的显微结构 Plate Ⅰ The microstructure of peripheral blood cells in three species 1,2,5. 青海沙蜥 Phrynocephalus vlangalii,3,4,7,8. 牛蛙 Rana catesbeiana,6. 中华蟾蜍 Bufo gargarizans; E. 红细胞 Erythrocyte,T. 血栓细胞 Thrombocyte,L. 淋巴细胞 Lymphocyte,M. 单核细胞 Monocyte,N. 嗜中性细胞 Neutrophil,B. 嗜碱性细胞 Basophil,A. 嗜酸/碱性细胞 Eosinophil/Basophil; 标尺 Bar=10 μm; 下图同,the same below. |
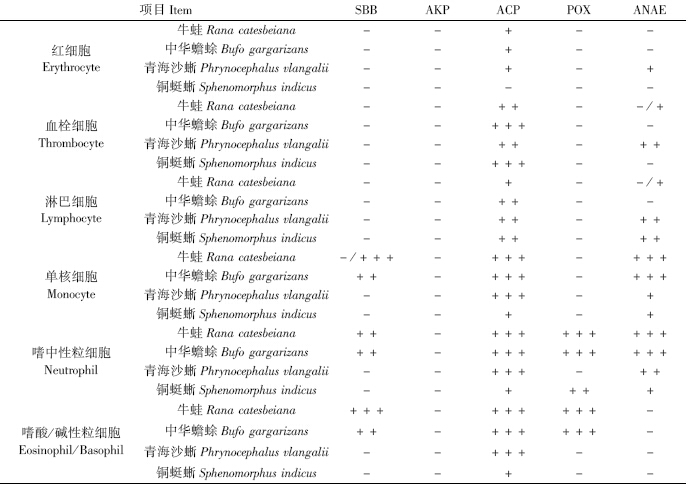
对4种动物外周血细胞进行细胞化学染色,结果见表 2。4种动物都具有嗜碱性粒细胞,但只有牛蛙、中华蟾蜍具有嗜酸性粒细胞。由于细胞化学染色很难区分嗜酸和嗜碱粒细胞,因此统称为嗜酸/碱性粒细胞。
|
牛蛙(图版Ⅱ:1)和中华蟾蜍(图版Ⅱ:2)的所有红细胞均含有少量ACP,呈颗粒[KG(2x]状或弥散状。青海沙蜥红细胞部分呈弱阳性(图版Ⅱ:3),其中阳性反应的红细胞比阴性的多。牛蛙和中华蟾蜍血栓细胞分别表现为阳性和强阳性(图版Ⅱ:4~5)。青海沙蜥和铜蜓蜥的血栓细胞则分别呈阳性和强阳性(图版Ⅱ:6~7)。牛蛙淋巴细胞有少量的ACP,呈弥散状分布(图版Ⅱ:8);中华蟾蜍淋巴细胞表现为阳性(图版Ⅱ:9)。青海沙蜥和铜蜓蜥的淋巴细胞均含有较多ACP(图版Ⅱ:10~11)。牛蛙和中华蟾蜍的单核细胞均表现为强阳性,胞质中充满红色团块状阳性反应物(图版Ⅱ:12~13);青海沙蜥和铜蜓蜥的单核细胞分别表现为强阳性和弱阳性(图版Ⅱ:14~15)。牛蛙和中华蟾蜍嗜中性粒细胞(图版Ⅱ:16)均呈强阳性;青海沙蜥和铜蜓蜥嗜中性粒细胞分别呈强阳性和弱阳性(图版Ⅱ:17~18)。牛蛙和中华蟾蜍的嗜酸/碱性粒细胞(图版Ⅱ:19~20)均呈强阳性。青海沙蜥和铜蜓蜥嗜碱性粒细胞分别呈强阳性(图版Ⅲ:21)和弱阳性(图版Ⅲ:22)。

|
| 图版Ⅱ4种两栖爬行动物外周血细胞的细胞化学特征(一) Plate Ⅱ The cytochemical characteristics of peripheral blood cells in four species 1~20. 酸性磷酸酶染色 Acid phosphatase(ACP)staining: 1,4,8,12,19. 牛蛙 Rana catesbeiana,2,5,9,13,16,20. 中华蟾蜍 Bufo gargarizans,3,6,10,14,17. 青海沙蜥Phrynocephalus vlangalii,7,11,15,18. 铜蜓蜥 Sphenomorphus indicus. |
中华蟾蜍单核细胞(图版Ⅲ:23)呈阳性,胞质中黑色阳性物弥散状分布,而牛蛙的部分单核细胞呈强阳性反应,其中强阳性反应的细胞(图版Ⅲ:24)比阴性细胞稍多。牛蛙和中华蟾蜍嗜中性粒细胞均呈阳性反应(图版Ⅲ:25~26)。牛蛙和中华蟾蜍的嗜酸/碱性粒细胞分别呈强阳性(图版Ⅲ:27)和阳性(图版Ⅲ:28)。

|
| 图版Ⅲ4种两栖爬行动物外周血细胞的细胞化学特征(二) Plate Ⅱ The cytochemical characteristics of peripheral blood cells in four species 21~22. 酸性磷酸酶染色Acid phosphatase staining: 21. 青海沙蜥Phrynocephalus vlangalii,22. 铜蜓蜥Sphenomorphus indicus; 23~28. 苏丹黑B染色Sudan black B staining: 23,26,28. 中华蟾蜍Bufo gargarizans,24,25,27. 牛蛙Rana catesbeiana; 29~33. 过氧化物酶反应Peroxidase staining: 29,32. 牛蛙R. catesbeiana,30,33. 中华蟾蜍B. gargarizans,31. 铜蜓蜥S. indicus; 34~45. 酸性-α-醋酸萘酯酶染色α-naphthyl acetate esterase staining: 34,36,38,42. 牛蛙R. catesbeiana,35,40,44. 青海沙蜥P. vlangalii,37,41,45. 铜蜓蜥S. indicus,39,43. 中华蟾蜍B. gargarizans; 标尺Bar=10 μm。 |
牛蛙和中华蟾蜍嗜中性粒细胞(图版Ⅲ:29~30)均呈强阳性;铜蜓蜥嗜中性粒细胞(图版Ⅲ:31)呈阳性反应。牛蛙和中华蟾蜍的嗜酸/碱性粒细胞(图版Ⅲ:32~33)均呈强阳性。 2.2.4 ANAE染色
牛蛙部分血栓细胞为弱阳性,其中阳性细胞(图版Ⅲ:34)比阴性细胞多。青海沙蜥血栓细胞则呈阳性(图版Ⅲ:35)。牛蛙部分淋巴细胞呈弱阳性反应,其中阳性反应的淋巴细胞(图版Ⅲ,36)比阴性细胞稍多。青海沙蜥和铜蜓蜥的淋巴细胞均呈阳性反应(图版Ⅲ:37)。牛蛙和中华蟾蜍的单核细胞均呈强阳性(图版Ⅲ:38~39),说明含有大量ANAE。青海沙蜥和铜蜓蜥的单核细胞则均呈弱阳性(图版Ⅲ:40~41)。牛蛙和中华蟾蜍嗜中性粒细胞(图版Ⅲ:42~43)均呈强阳性。青海沙蜥和铜蜓蜥嗜中性粒细胞(图版Ⅲ:44~45)分别为阳性和弱阳性。4种动物的嗜碱性粒细胞均呈阴性。 3 讨论 3.1 红细胞的特点
红细胞的主要功能是运输氧气和二氧化碳。红细胞运输氧气的功能主要由血红蛋白来完成。一般而言,脊椎动物进化程度越高的种类,红细胞体积越小,数量越多,运输氧气的能力越强(郭宪光等,2002)。牛蛙和中华蟾蜍红细胞比青海沙蜥和铜蜓蜥的红细胞大。两栖动物比大部分爬行动物的红细胞大,而爬行动物的红细胞比哺乳动物的(人类成熟红细胞直径7 μm)大,这符合进化规律。牛蛙和中华蟾蜍与新疆北鲵Ranodon sibiricus(26~37 μm)(侯水薇等,1995)、大鲵Andrias davidianus(40~51 μm)(李丕鹏等,1989)等相比较,红细胞又较小,从一个方面说明无尾目比有尾目的进化地位要高。龟鳖类比蜥蜴类进化地位低,龟鳖类的红细胞较其他爬行类的大(吴孝兵等,1998)。 3.2 细胞化学特征
POX可降解过氧化物及过氧化脂质,消除对生物膜造成的损伤。随吞噬作用的开始细胞内会产生大量H2O2。而髓过氧化物酶可对H2O2和卤化物起作用,产生活性卤化物起杀菌作用(成令忠等,2003)。牛蛙和中华蟾蜍的粒细胞均含有POX,青海沙蜥外周血细胞均不含POX,而铜蜓蜥仅嗜中性粒细胞中含有POX。青海沙蜥POX的缺乏可能会直接影响其吞噬异物和杀菌的功能。青海沙蜥与鱼类中的胭脂鱼(金丽等,2011)、草鱼(郭琼林,卢全章,1994)、长薄鳅(黄小铭等,2012)相同,外周血细胞均缺乏POX;而南方鲇(张锡林等,1998)、长吻鮠(金丽等,2010)等鱼类嗜中性粒细胞中含有大量POX,可见,外周血细胞中POX的有无可能与种属差异有关,与进化地位无明显相关性。
ACP为溶酶体酶,是细胞吞噬、杀菌等功能的一种具有代表性的标志酶(Vazquez & Guerreero,2007)。牛蛙、中华蟾蜍、青海沙蜥和铜蜓蜥外周血细胞中ACP分布很广泛,所有白细胞均含ACP。这说明ACP是这4种动物外周血细胞抵抗病原体和异物入侵的主要酶类,外周血细胞中ACP的广泛分布或许可以在一定程度上弥补其POX的缺乏。
ANAE是哺乳类T淋巴细胞的细胞化学标志酶之一,在免疫防御中起作用。青海沙蜥外周血白细胞中除嗜碱性粒细胞之外均含有ANAE;铜蜓蜥的单核细胞、淋巴细胞和嗜中性粒细胞也均含有ANAE。中华蟾蜍的嗜中性粒细胞和单核细胞均含有ANAE;牛蛙的嗜中性粒细胞和单核细胞均含有ANAE,但淋巴细胞和血栓细胞仅部分含有ANAE。这种特点也体现了两栖类淋巴细胞存在分化差异。血栓细胞对非特异性酯酶反应的不同是否体现了其处于不同的功能状态,还不能确定。ANAE是哺乳类T淋巴细胞的标志酶,但在低等脊椎动物,如鱼类、两栖类、爬行类的外周血血栓细胞、单核细胞及嗜中性粒细胞中均不同程度地存在这种酶。海龟Chrysemys dorbignih单核细胞的ANAE与胞内毒素、小分子物质加工处理和抗原呈递有关(Azevedo & Lunardi,2003)。是否低等脊椎动物血栓细胞、单核细胞及嗜中性粒细胞中的ANAE均与胞内毒素、小分子物质加工处理和抗原呈递有关,还有待深入研究。
爬行类的新陈代谢高,主动适应环境的能力强于两栖类,而两栖类新陈代谢低,对环境的主动适应能力差,因此其相应的细胞类型和各种酶类也比较多,以应对各种不良环境。爬行类青海沙蜥各种酶类分布的外周血细胞类型比铜蜓蜥多,这可能与青海沙蜥的生活环境恶劣有关。
| 成令忠, 钟翠平, 蔡文琴. 2003. 现代组织学[M]. 上海: 上海科学技术文献出版社: 281-298. |
| 龚志锦, 詹镕洲, 董荣春. 1994. 病理组织制片和染色技术[M]. 上海: 上海科学技术文献出版社: 138-139, 313-317. |
| 郭琼林, 卢全章. 1994. 草鱼肾脏、脾脏血细胞发育过程超微结构与细胞化学的研究[J]. 水生生物学报, 18(3): 240-246. |
| 郭宪光, 张耀光, 王志坚, 等. 2002. 中华蟾蜍血细胞的观察[J]. 四川动物, 21(4): 211-214. |
| 侯水薇, 王秀玲, 王志勇, 等. 1995. 新疆北鲵血液的组织学观察[J]. 两栖爬行动物学研究, (4, 5辑): 58-62. |
| 胡知渊, 来雅萍, 陈文静. 2005. 棘胸蛙、虎纹蛙、黑斑蛙血细胞的比较[J]. 四川动物, 24(1): 5-9. |
| 黄斌, 杨霁虹, 陆玉建. 2002. 巴西彩龟血细胞显微结构的观察研究[J]. 动物科学与动物医学, 19(9): 16-18. |
| 黄小铭, 张耀光, 江星, 等. 2012. 长薄鳅外周血细胞的显微结构和细胞化学特征研究[J]. 四川动物, 31(1): 59-63. |
| 金丽, 杜长雷, 张耀光. 2010. 长吻鮠血细胞的显微、亚显微结构及细胞化学研究[J]. 西南大学学报: 自然科学版, 32(12): 79-85. |
| 金丽, 赵娜, 黄林, 等. 2011. 胭脂鱼外周血细胞的显微、超微结构与细胞化学观察[J]. 水生生物学报, 35(3): 550-556. |
| 李桂芬, 蒙绍权, 李涛, 等. 2009. 版纳鱼螈外周血细胞观察[J]. 动物学杂志, 44(2): 102-107. |
| 李丕鹏, 何国湘, 张育辉, 等. 1989. 大鲵的血液学观察[J]. 陕西师范大学学报: 自然科学版, 17(3): 50-53. |
| 刘志洁, 宗英, 张金国, 等. 2002. 野生动物血液细胞学图谱[M]. 北京: 科学出版社: 2-4. |
| 王石泉. 1995. 鳖外周血细胞显微形态及细胞化学[J]. 动物学杂志, 30(1): 16-19. |
| 吴孝兵, 张盛周, 吴海龙, 等. 1998. 16种爬行动物血细胞形态学参数研究[J]. 动物学杂志, 33(1): 29-31. |
| 张锡林, 王明清, 蒋灵芝. 1998. 大口鲶外周血细胞的结构和组化反应的研究[J]. 四川动物, 17(3): 100-101. |
| 周庆萍, 李松, 黄倩. 2010. 双团棘胸蛙血细胞的显微观察[J]. 西北师范大学学报: 自然科学版, 46(4): 87-90. |
| Azevedo A, Lunardi LO. 2003. Cytochemical characterization of eosinophlic leukocytes circulating in the blood of turtle (Chrysemys dorbignih)[J]. Acta Histochem, 105(1): 99-105. |
| Vazquez GR, Guerrero GA. 2007. Characterization of blood cells and hematological parameters in Cichlasoma dimerus (Teleostei, Perciformes)[J]. Tissue and Cell, 39: 151-160. |
| Tavares-Dias M. 2006. A morphological and cytochemical study of erythrocytes, thrombocytes and leukocytes in four freshwater teleosts[J]. J Fish Biol, 68(6): 1822-1833. |