 2015, Vol. 34(2)
2015, Vol. 34(2)扩展功能
文章信息
- 刘睿, 毕俊怀, 付明霞, 陈绍勇
- LIU Rui, BI Junhuai, FU Mingxia, CHEN Shaoyong
- 鄂尔多斯地区密点麻蜥的食性分析
- The Diet of Eremias multiocellata in Erdos, China
- 四川动物, 2015, 34(2): 193-199
- Sichuan Journal of Zoology, 2015, 34(2): 193-199
- 10.3969/j.issn.1000-7083.2015.02.006
-
文章历史
- 收稿日期:2014-06-04
- 接受日期:2014-11-17






食物是联系动物与环境的纽带,食性一方面决定了动物在生物群落中的生态位(Goodman,1871),另一方面影响动物营养水平和栖息地结构(Germano et al.,2007)。动物的食性研究一直是生态学中一项主要的研究内容。同时,动物的种群动态、行为、生理等均有赖于食性研究。实践中,确定动物的食性也是进行种群科学管理的前提(吴建平,2007)。密点麻蜥Eremias multicellata是我国北部和西北部荒漠草原和半荒漠草原最常见的一种卵胎生蜥蜴,在生态系统中属于消费者,保证能量遵循食物链和食物网逐级流动,在荒漠化草原生态系统中起着举足轻重的作用。密点麻蜥食性很广,捕食大量有害昆虫,对消灭荒漠害虫,保护沙生植物有重要意义(毕俊怀等,1997)。近年来,在蜥蜴食性分析方面已经有一些研究报道(刘迺发,李仁德,1995;刘迺发,耿稚荣,1995;Christian et al.,1996;孙建梅,2000;赵文阁等,2006)。连雪(2011)对草原沙蜥食物多样性及营养生态位宽度做了初步统计并对其有益系数做了计算,但样本量较少。孙建梅(2000)的研究中只提出了山地麻蜥的食物以农林害虫为主,并未对有益系数进行计算。因此对密点麻蜥的食性做深入研究是十分必要的。本研究尝试用较大的样本量,深入研究了鄂尔多斯库布齐沙漠不同年龄和不同性别密点麻蜥食性的差异及其有益系数。 1 材料与方法 1.1 自然地概况
密点麻蜥在内蒙古自西向东的分布区域依次为浑善达克沙地、库布齐沙漠、毛乌素沙地、乌兰布和沙漠、腾格里沙漠和巴丹吉林沙漠,除此之外在干旱草原以及退化的草原上也有分布。本次试验的密点麻蜥于2012年8—9月于鄂尔多斯库布齐沙漠(40.213°N,111.115°E;1042.4 m)采集,该地属于典型荒漠草原生态系统,靠近黄河由北向南流的部分。 1.2 实验动物采集与保存
本研究于2012年8—9月10∶00—16∶00蜥蜴活动高峰期捕捉(陈小勇等,1997),在尽量不影响种群动态及长期食性变化的范围内捕捉,捕获后立即注射75%乙醇于蜥蜴腹腔内处死并浸于75%乙醇保存液中带回实验室备检。[=DM(]刘睿等: 鄂尔多斯地区密点麻蜥的食性分析 1.3 食性分析
将带回的野外标本解剖观察,获得动物种群短期的食性,分析不同年龄和性别密点麻蜥的食性,并为密点麻蜥的益害性提供有效评价。通过剖胃法对不同性别和年龄组成的样本进行食性研究。取出整胃,在SMZ168型解剖镜下进行解剖鉴定,统计胃内食物种类及数量。对于不完整个体及半消化状态的食物,依残存的昆虫复眼、触角、口器、翅、颚、附肢及腹板等进行鉴定。参照《昆虫学》(南开大学等,1980)、《内蒙古昆虫》(能乃扎布,1999)、《昆虫分类学》(蔡邦华,1973)和《昆虫分类检索表》(李鸿兴等,1987)进行鉴定,鉴定过程参考Tatner(1983)提供的方法。本研究共分析85个胃的内容物,包括雄性20个、雌性56个,其中,成体59个、亚成体17个、幼体9个。 1.4 数据统计
食物数量百分比K=该类食物在胃内出现的数量/各食物类群在胃内出现的总数量×100%。
食物频次百分比P=该类食物在胃内出现的频次/各食物类群在胃内出现的总频次×100%。
食物百分率相似性指数(index of percent similarity,PS)PS=Σ(Pmin),Pmin为每一科最低的食物数量百分比。
食物百分率相异性指数(index of percent diversity,PD)(孙儒泳,2001)PD=1-PS。
营养生态位重叠(overlap of trophic ecological niche,O)O=1-1/Σ|Pij-Pjk|。Pij、Pjk分别为被j和k动物组利用的第i种食物在所有被利用的食物中所占的频次百分比。
营养生态位分离值(separation of trophic ecological niche,S)S=1-O。
营养生态位宽度(width of trophic ecological niche,B)B=1/∑P2i。Pi为实际利用的第i种食物在所有被利用的食物中所占的频次百分比。食物多样性指数和营养生态位宽度呈正相关,多样性指数越高,摄食种类越多,越均匀,营养生态位越宽,摄食越广泛(刘迺发,李仁德,1995)。
食物类群的丰富度(Shannon-Wiener index,H)H=-∑(Pi)(log2Pi),式中,Pi为第i类群在胃中的食物数量百分比(孙儒泳,2001)。
竞争指数(competition coefficient,a)aj(k)=∑(PikPij)/∑Pij2。
摄食率=含食物的胃数/总胃数×100%。
有益系数V=(n-u)/t,其中,n为有害动物个体总数,u为有益动物个体总数,t为动物个体总数。计算时,益害兼有及益害不明两类的数据不统计在有益或有害的任一类中,但在计算为内容物总数(t)时,则包含4种类型的数据总和(梁中宇等,1958)。 2 结果 2.1 密点麻蜥胃容物组成
鄂尔多斯库布齐沙漠密点麻蜥食谱较广,经鉴定共有动物和植物2类。本研究对85个胃的内容物分析结果为:空胃5个,实胃80个,摄食率94.12%,其中雌性摄食率92.86%、雄性摄食率95.00%。共鉴定出2纲13目41科的动物性食物。取食食物共计1088个,在部分胃容物中发现植物。在各食物类群中,按食物数量、食物数量百分比、出现频次和频次百分比的综合判断,密点麻蜥以动物性食物为主,植物次之,为杂食性动物。通过食物数量百分比和遇见率综合判断,其中密点麻蜥最喜爱的食物种类包括蚜科Aphididae(54.69%、62.34%)、鳞翅目Lepidoptera幼虫(6.89%、48.24%)、尺蛾科Geometridae(3.03%、25.88%)和蜘蛛目Araneida(2.85%、23.53%)(表 1)。
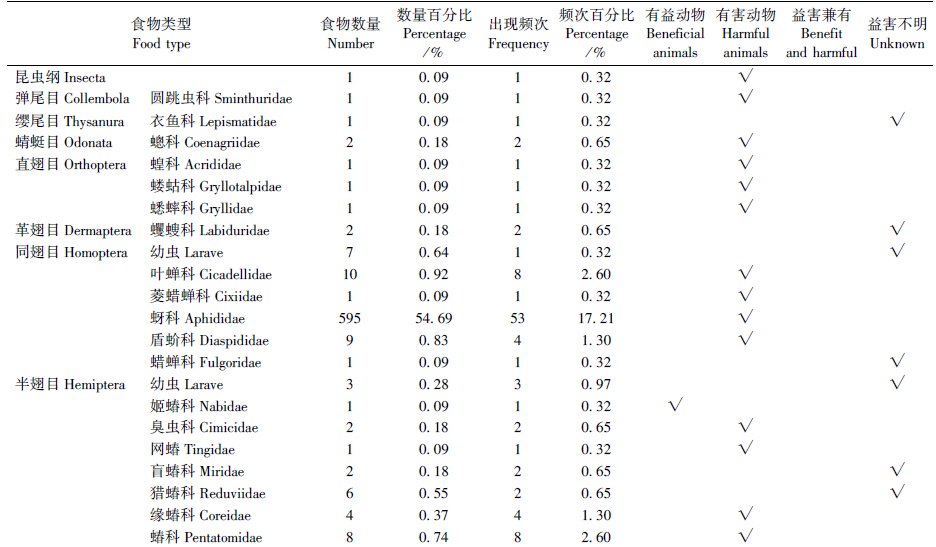
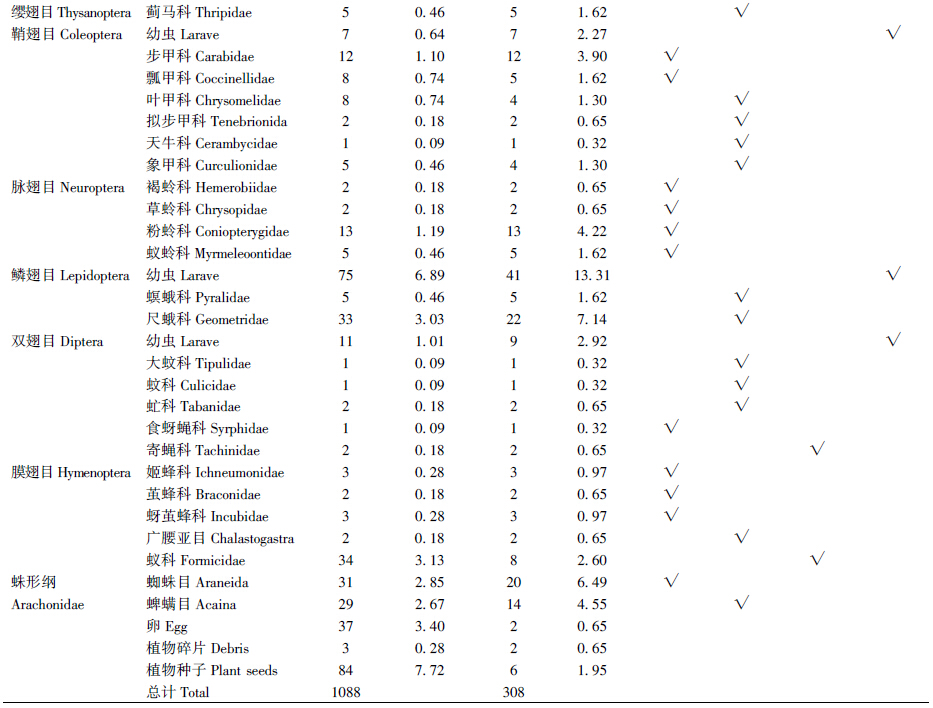
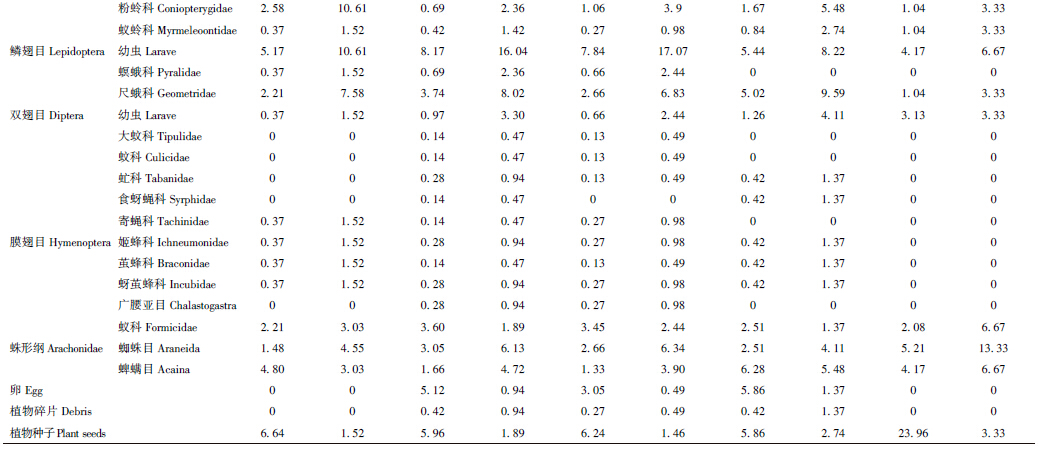
通过对食物数量百分比、频次百分比的综合判断,密点麻蜥雌、雄两性的主要食物都是蚜科(雄性63.84%、21.21%,雌性53.74%、16.04%)和鳞翅目幼虫(雄性5.17%、10.61%,雌性8.17%、16.04%)(表 2)。次要食物在两性间不同,雄性是粉蛉科(2.58%、10.61%),而雌性则是尺蛾科(3.74%、8.02%)。密点麻蜥雌、雄个体的食物百分率相似性指数(78.96)及营养生态位重叠值(0.98)均较大,食物竞争系数(0.97)、食物百分率相异性指数(21.04)及营养生态位分离值(0.02)均较小,说明雌、雄个体之间的食物相似程度大,食物竞争强烈。雄性密点麻蜥的食物多样性指数(2.34)及营养生态位宽度(11.22)均小于雌性(分别为2.98和13.72)(表 3)。

|
|

|
通过对食物数量百分比、频次百分比的综合判断,密点麻蜥成体的主要食物是蚜科(59.50%、18.05%)和鳞翅目幼虫(7.84%、17.07%),次要食物是尺蛾科(2.66%、6.83%)。亚成体的主要食物是蚜科(48.11%、17.80%)和尺蛾科(5.02%、9.59%),次要食物是鳞翅目幼虫(17.07%、5.44%)。幼体的主要食物是蚜科(35.42%、16.67%),次要食物是蜘蛛目(5.21%、13.33%)。
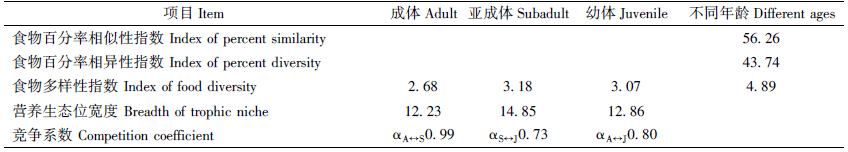
不同年龄密点麻蜥间的食物百分率相似性指数值较小(56.26),食物百分率相异性指数为43.74,说明不同年龄间密点麻蜥的取食种类略有差异。不同年龄个体间食物相似程度不同,食物竞争强烈程度均不同。成体与亚成体间的食物竞争系数α(αAS0.99),亚成体与幼体间的食物竞争系数α(αSJ0.73),成体与幼体间的食物竞争系数α(αAJ0.80)。不同年龄的密点麻蜥食物多样性指数及营养生态位宽度均为亚成体最大(3.18和14.85),成体最小(2.68和12.23)(表 4)。
|
通过表 1数据对密点麻蜥进行益害评价,运用有益系数公式计算密点麻蜥总有益系数,V=59.47%,V为正数且数值较大,说明密点麻蜥取食害虫多于益虫。 3 讨论
食性研究是从营养学的角度探讨动物取食策略、生境适应机制的基础(吴建平等,2007)。本次研究采集样本胃容物共鉴定2纲13目41科动物物种,食物数量与食物出现频率最高的均为同翅目蚜科。鲍敏等(1998)发现密点麻蜥多生活在海拔3500 m以下的高原、丘陵和盆地的干旱草原上及荒漠、半荒漠边缘的稀疏灌丛地带,主要以甲虫、蚂蚁、蛾、蝇、蚊等昆虫为食,同时也以植物性食物白刺果和野枸杞果为食。而鄂尔多斯库布齐沙漠地区密点麻蜥生活于半干旱地带,风蚀沙化现象十分严重,属于典型的荒漠草原生态系统,海拔1100 m,植被分布具有地带性。这表明密点麻蜥的食性与栖息地生境可能有直接关系。所食昆虫种类具有明显的季节变化,这与昆虫的季节性发生、活动及植物生长有关。此外,麻蜥的食性可能存在种间差异。冯照军等(2005)对山地麻蜥E. brenchleyi进行研究,结果表明食物数量与食物出现频率最高的均为膜翅目昆虫。赵德义等(2000)对丽斑麻蜥E. argus进行研究,结果表明其食物以膜翅目、鞘翅目、双翅目为主。
最优化觅食理论认为动物在觅食过程中搜寻食物和进食过程都需要花费能量,从食物中获得的能量与觅食过程中的能量消耗之差即为动物摄食的净收入,摄取某种食物所得收入越大,这种食物对捕食者来说就越有利,因此捕食者在觅食过程中总是选择有利性更大的食物(王培潮,1964)。在胎生蜥蜴的食谱中,同翅目蚜科、蛛形纲动物和鳞翅目幼虫所占比例较大,这与这些动物相对容易捕食、易消化且具有较高的能量有关,捕食这些动物可以使胎生蜥蜴获得更大的净收入。
两性之间,雌性密点麻蜥食物生态位宽度与食物多样性指数均较大且成正比,这可能与繁殖有关。经分析,雌、雄两性成体食物生态位重叠度较大,但它们通过不同的觅食对策进行适应(宋志明,赵肯堂,1983),两性个体存在强烈的竞争可能性,但由于不同年龄个体食物百分率相似性系数较小,这样生态位的分离一定程度上可以缓解种内食物资源的竞争。
食性在年龄和性别上的差异可以保证动物在不同发育阶段上对营养的不同需要,同时又扩大了该种动物与环境食物的联系,因而在更大范围内利用环境条件,对促进物种的分布范围扩大和个体数量增加很有意义(孙儒泳,1992)。不同年龄密点麻蜥的食性有不同程度的差异,亚成体的营养生态位宽度与食物多样性指数最大,而成体最小。这可能与亚成体性腺与身体的逐渐发育和增大,采用不同觅食对策,以满足生长发育与生殖活动的需要有关。幼体食物较小并且在觅食过程中更多选择高能量的食物。这可能与幼体个体较小,难以吞食较大、较硬的食物且高能量食物能够满足快速生长发育有关。群落成员之间在体形大小方面通常存在着有规律的差异,体形比例最小的个体必然会减少竞争,而保持稳定共存(Bowers & Brown,1982)。
通过计算密点麻蜥动物性食物组成的益害关系,总有益系数为正数且数值较大,说明密点麻蜥取食害虫数目比益虫多,是对荒漠化草原有益的类群,对生态系统中的虫害有一定控制作用。
致谢:感谢能乃扎布教授百忙中对昆虫的鉴定,同时感谢中国科学院动物研究所师生所给予的支持与帮助。
| 鲍敏, 曾阳, 马建滨, 等. 1998. 青海沙蜥和密点麻蜥的分布状况、生活规律及食性[J]. 青海师范大学学报: 自然科学版, 4: 42-45. |
| 毕俊怀, 李建军, 苏兆龙. 1997. 内蒙古三种蜥蜴的食性研究[M]// 李德俊. 两栖爬行动物学研究(6-7). 贵阳: 贵州科技出版社: 123-127. |
| 蔡邦华. 1973. 昆虫分类学: 中册[M]. 北京: 科学出版社: 8-100. |
| 陈小勇, 王建平, 龚大洁. 1997. 兰州北山三种蜥蜴春季食性分析[J]. 动物学杂志, 32(5): 13-18. |
| 冯照军, 孙建梅, 赵彦禹, 等. 2005. 山地麻蜥生态的初步研究[J]. 四川动物, 24(3): 359-366. |
| 李鸿兴, 隋敬之, 周世秀, 等. 1987. 昆虫分类检索表[M]. 北京: 农业出版社. |
| 连雪. 2011. 草原沙蜥(Phrynocephalus frontalis)种群的时空分布、栖息地选择及其对控制虫害的作用[D]. 呼和浩特: 内蒙古师范大学. |
| 梁中宇, 刘良材, 吴其荣. 1958. 秋收时几种稻田蛙胃内容物的初步分析[J]. 动物学杂志, 2(4): 220-229. |
| 刘逎发, 耿稚荣. 1995. 密点麻蜥的食性及摄食的季节性变化[J]. 应用生态学报, 6: 74-78. |
| 刘逎发, 李仁德. 1995. 三种荒漠蜥蜴空间和营养生态位研究[J]. 生态学报, 15(1): 48-53. |
| 南开大学, 中山大学, 北京大学, 等. 1980. 昆虫学: 上册[M]. 北京: 高等教育出版社: 106-138. |
| 能乃扎布. 1999. 内蒙古昆虫[M]. 呼和浩特: 内蒙古人民出版社. |
| 宋志明, 赵肯堂. 1983. 草原沙蜥和密点麻蜥的食性研究[J]. 两栖爬行动物学报, 2(4): 7-12. |
| 孙建梅. 2000. 山地麻蜥春秋季食性的研究[J]. 徐州师范大学学报: 自然科学版, 18(4): 65-66. |
| 孙儒泳. 1992. 动物生态学原理[M]. 北京: 北京师范大学出版社. |
| 孙儒泳. 2001. 动物生态学原理[M]. 北京: 北京师范大学出版社. |
| 王培潮. 1964. 杭州四种蜥蜴的生态研究I. 分布状况、活动规律及食性[J]. 动物学杂志, 6(2): 70-76. |
| 吴建平. 2007. 小兴安岭地区原麝冬季食性研究[J]. 兽类学报, 1: 58-63. |
| 赵德义, 丁淑萍, 曾桂荣. 2000. 丽斑麻蜥的生态学研究[J]. 高师理科学刊, 20(2): 52-53. |
| 赵文阁, 董丙君, 刘鹏, 等. 2006. 胎生蜥蜴的捕食行为及其食性分析[J]. 生态学杂志, 25(12): 1520-1523. |
| Bowers MA, Brown JH. 1982. Body size and coexistence in desert rodents: chance or community structure[J]. Ecology, 63: 391-400. |
| Christian KA, Griffiths AD, Bedford GS. 1996. Physiological ecology of frillneck lizard in a seasonal tropical environment[J]. Oecologia, 106(1): 49-56. |
| Germano DJ, Smith PT, Tabor SP. 2007. Food habits of the blunt-nosed leopard lizard(Gambelia sila)[J]. The Southwestern Naturalist, 52(2): 318-323. |
| Goodman N. 1871. The descent of man and selection in relation to sex[J]. Journal of Anatomy and Physiology, 5(2): 363-372. |
| Tatner P. 1983. The diet of urban magpies Pica pica[J]. Ibis, 125: 90-107. |