 2015, Vol. 34(2)
2015, Vol. 34(2)扩展功能
文章信息
- 戴波, 陈本平, 岳碧松, 曾涛
- DAI Bo, CHEN Benping, YUE Bisong, ZENG Tao
- 四川山鹧鸪栖息地破碎化及保护管理状况分析
- A Study on Habitat Fragmentation and Conservation Status of Arborophila rufipectus
- 四川动物, 2015, 34(2): 174-180
- Sichuan Journal of Zoology, 2015, 34(2): 174-180
- 10.3969/j.issn.1000-7083.2015.02.003
-
文章历史
- 收稿日期:2014-06-30
- 接受日期:2014-10-17






2. 四川大学 生命科学学院, 四川省濒危野生动物保护生物学重点实验室, 生物资源与生态环境教育部重点实验室, 成都 610064;
3. 四川老君山国家级自然保护区管理局, 四川屏山 645350
2. Sichuan Key Laboratory of Conservation Biology on Endangered Wildlife, Key Laboratory of Bio-resources and Eco-environment of Ministry Education, College of Life Sciences, Sichuan University, Chengdu 610064, China;
3. Sichuan Laojunshan National Nature Reserve Management Bureau, Pingshan, Sichuan Province 645350, China
四川山鹧鸪Arborophila rufipectus是我国特有珍稀鸟类,目前所知仅见于四川省中南部和云南省东北部,分布区域狭窄,数量稀少,各种自然和人为因素的影响使四川山鹧鸪生存受到威胁,亟待保护,其濒危程度已引起国内外广泛关注。我国政府将其定为国家Ⅰ级重点保护野生动物,世界自然保护联盟(IUCN)在其编制的《世界濒危物种红皮书》中将其列为濒危(EN)等级(IUCN,2014),IUCN物种生存委员会、国际鸟类联合会(Bird Life International)和世界雉类协会(WPA)鹑类专家组将四川山鹧鸪及其栖息地保护管理作为鹑类保护行动计划最优先的领域。
四川山鹧鸪生活隐秘,深入研究困难,近年的研究工作主要集中在分布数量(Dai et al.,1998;李操等,2003;陆钢等,2007;何芬奇等,2008,2009)、生境利用(李操等,2003;Liao et al.,2007,2008a,2008b;Dai et al.,2009)、繁殖行为和生态(廖文波等,2007;廖文波,胡锦矗,2010)、线粒体DNA测序及相关物种系统发育(He et al.,2009)等方面,并取得了一些进展,获得的相关成果资料为该物种的保护管理奠定了基础。但到目前为止,我们对四川山鹧鸪的认知尚且肤浅,所得信息远不能满足科学有效管理的需要。了解栖息地景观特征是物种保护宏观决策的前提。戴波等(2014)从景观尺度对四川山鹧鸪的分布格局和状况做了初步研究,但未涉及详细的栖息地景观特征。本文在其基础上定量分析了该物种栖息地景观特征,结合栖息地保护现状,探讨保护管理策略。 1 材料与方法 1.1 遥感数据处理
遥感数据采用美国马里兰大学"全球地被机构"(Global L and Cover Facility,GLCF)数据库中保存的6幅涵盖四川山鹧鸪已知分布区及周边区域的L and Sat 7 ETM+卫星影像,分别为2001年6月14日3幅、2001年4月18日2幅和2002年4月21日1幅。采用戴波等(2014)所用遥感数据处理方法和模型解译,得出四川山鹧鸪栖息地分布矢量图层。整个过程在ArcView 3.3、ArcGIS 10.0和ERDAS Imagine 9.2软件平台中完成。该栖息地分布图层的准确性和有效性对于景观尺度上做四川山鹧鸪栖息地评估是适合的(戴波等,2014)。 1.2 栖息地破碎化分析
从斑块和景观(包括分类景观和总体景观)两个层次,选取与景观破碎化密切相关的斑块量度、边缘状况、形状特征三方面的参数,分析四川山鹧鸪栖息地的破碎化程度。在ArcGIS软件中,载入上述四川山鹧鸪栖息地分布矢量图层,采用"斑块分析"扩展模块(Rempel et al.,2012)做相关运算:在斑块层面,分别求算图层中每个斑块的周长面积比(Edge Density)、形状指数(Shape Index)、分维度(Fractal Dimension)。在景观层面,对现实栖息地、潜在栖息地和栖息地总体,计算斑块数量、平均大小、边缘密度、平均边缘长度、平均形状指数、面积加权平均形状指数、平均分维度和面积加权平均分维度。由此获得栖息地破碎化程度的特征信息;另用ArcGIS"邻域工具",筛选与四川山鹧鸪栖息地各斑块的最邻近斑块,并计算边缘之间的距离,再找出与各斑块最邻近的现实栖息地斑块,计算边缘上最近点之间的距离,以了解斑块之间的邻接程度和连通的可能性。 1.3 栖息地保护管理状况分析
根据四川、云南自然保护区主管部门最新发布的自然保护区信息,得到四川山鹧鸪分布区的自然保护区名称、类型、级别、面积等建设现状,进一步从各保护区所在地政府部门了解具体位置信息,生成保护区分布图层,与上述四川山鹧鸪栖息地分布图叠加,从区位相关性了解栖息地受保护现状。 2 结果与分析 2.1 四川山鹧鸪栖息地破碎化程度
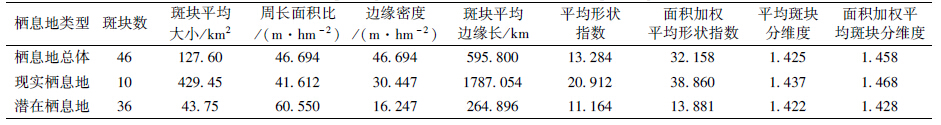
四川山鹧鸪栖息地的斑块分析结果见表 1和表 2。
|

|
理论上,景观包含的斑块数越少、平均面积越大,景观完好程度越高。多边形边界越简单、平滑,其边缘长度和密度越低。多边形外形越规则、形状越正,其形状指数和分维度越小。正圆形的边缘长度和密度最低,形状指数和分维度均为1,而边界越简单、外形越接近圆形的多边形,其形状指数和分维度越趋于1(McGarigal & Marks,1994)。表 1显示,四川山鹧鸪栖息地整体构成多达46个斑块,被割裂的程度可见一斑。此外,无论是总体上看,还是按现实栖息地和潜在栖息地分类来考量,栖息地斑块的平均边缘长度和边缘密度都比较高,平均形状指数较大,平均斑块分维度偏离1多达0.4以上,这些均说明其栖息地边缘复杂,形状不规整,呈现很高的破碎化程度。值得注意的是,相对于潜在栖息地而言,现实栖息地的边缘密度、平均边缘长度、平均形状指数和平均分维度都更高,尤其是边缘密度、平均形状指数几乎超出潜在栖息地的1倍,平均边缘长度超出1个数量级,面积加权平均形状指数高出2倍,所以现实栖息地的空间外形更为不规则,边缘复杂程度更高,更容易被分割而进一步破碎化。另一方面,潜在栖息地斑块数远大于现实栖息地,占总块数的78%,但其斑块平均大小则仅约为后者的10%,远低于总体平均值,因此潜在栖息地相对很零碎,而且其周长面积比高于现实栖息地和总体平均值,说明其更易于受外界干扰影响,不容忽视。
如表 2所示,就单个栖息地斑块来看,现实栖息地中面积最大的2片,即"甘洛-金口河-峨边-马边-美姑"片(XS01)和"雷波-马边-屏山"片(XS03)的周长面积比较小,不到40 m·hm-2,理论上抗外部干扰的能力较强。相反,"甘洛东a"片(XS02)和"雷波中南a"片(XS08)周长面积比较大,超出60 m·hm-2,相对更易受到外部干扰影响;而从形状指数和分维度看,"甘洛a"片(XS02)和"田坝-大红岩"片(XS06)较小,因此这2片栖息地的空间外形相对较圆满,完整程度较好。相对的是"甘洛-金口河-峨边-马边-美姑"片(XS01)和"绥江-水富-永善-盐津-大观"片(XS03)的形状指数和分维度较大,其外形相对更加不规整,易于被分割断裂。
对于潜在栖息地,"大风顶北段"片(QZ11)周长面积比最低,其次是"瓦山"片(QZ03)和"镇雄d"片(QZ36),表明它们边缘相对平滑。而"甘洛南b"片(QZ08)、"大关南a"片(QZ22)、"彝良东北"片(QZ28)、"彝良东南"片(QZ30)的周长面积比大,超过100 m·hm-2,它们相对更易受到外部干扰影响;形状指数和分维度数据表明"镇雄西d"片(QZ36)形状最规整,其次是"永善北b"片(QZ16)及"镇雄西a"片(QZ33)。而"甘洛南a"片(QZ07)、"永善北a"片(QZ15)、"大关东-盐津南"片(QZ21)形状相对复杂,更易破碎化。 2.2 四川山鹧鸪栖息地邻接程度
表 2中也列出了邻域分析结果。结果表明,四川山鹧鸪部分栖息地斑块非常接近,如"雷波-马边-屏山"片(XS03)与"大风顶北段"片(QZ11)、"甘洛-金口河-峨边-马边-美姑"片(XS01)与"瓦黑北"片(QZ10)、"甘洛-金口河-峨边-马边-美姑"片(XS01)与"大风顶北段"片(QZ11)、"雷波中南a"片(XS08)与"雷波中南b"片(XS09)、"甘洛南a"片(QZ07)与"甘洛南b"片(QZ08)、"永善北"b(QZ16)片与"永善东-大关西a"片(QZ17)、"甘洛东b"(QZ06)片与"甘洛-金口河-峨边-马边-美姑"片(XS01),最近点距离都不到30 m。如此小的间距,是解译栅格图转换为矢量图层时斑块边界重建过程算法误差造成的,从原始卫片数据30 m的空间分辨率来讲,实际上应该为约1个方形像素边长或对角线长度的距离,即30~42.4 m。其他一些较为相接近的片区有:"箐口山"片(QZ13)与"雷波-马边-屏山"片(XS03)、"黄连山-五指山北段"片(XS04)与"老君山-五指山南段"片(XS05)、"永善北a"(QZ15)片与"永善北b"(QZ16)片,它们两两间最近点距离约2个像素宽。根据实地调查,这些对应斑块之间主要为以往林木采伐和交通运输修建的公路相隔(戴波等,2014),斑块之间在局部地段很可能存在一定的联系,如果公路沿线的植被得到保护和恢复,是可以连接起来的。
从各斑块最邻近的现实栖息地斑块指向情况看,"绥江-永善-水富-盐津-大关"片区(XS10)频次最高,它被多达23个栖息地斑块确定为最邻近的现实栖息地斑块。其次是"甘洛-金口河-峨边-马边-美姑"片(XS01),有9个栖息地斑块将其确定为最邻近的现实栖息地斑块。再次是"雷波-马边-屏山"片(XS03),有5个栖息地斑块确定其为最邻近的现实栖息地斑块。因此,这3片起着凝聚其他栖息地斑块的重要作用,加之它们是面积排前三的现实栖息地,整体上确实应为四川山鹧鸪的核心分布地带。 2.3 四川山鹧鸪栖息地保护管理状况
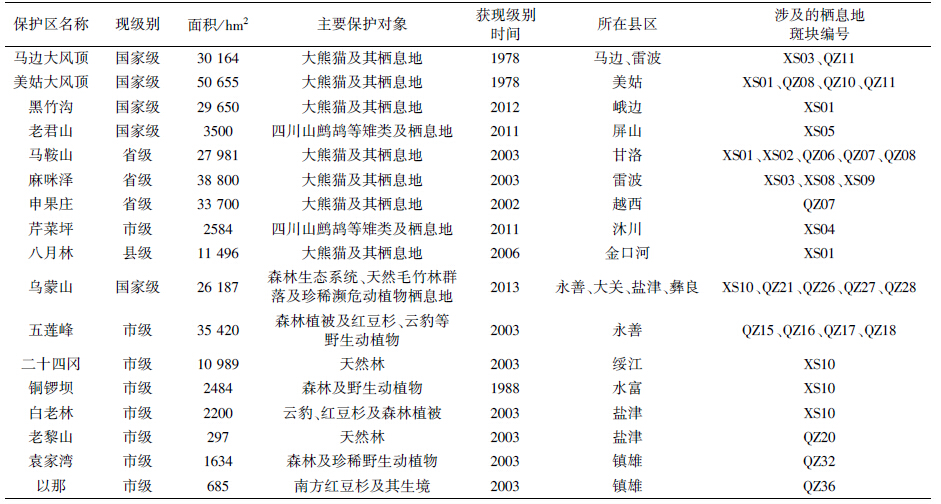
图 1显示了研究区内已建立的森林及陆生野生动物类型自然保护区。表 3列出了这些保护区现状及与四川山鹧鸪栖息地的关系。可知目前四川山鹧鸪分布区内已建保护区17个,总面积308 426 hm2。其中国家级5个,省级3个,市级8个,县级1个。只有2个是20世纪70年代建立的老保护区,其他均为20世纪90年代或21世纪初新建。这些保护区初步形成了四川山鹧鸪分布区内的保护网络,其管护范围涉及24个栖息地斑块,占总斑块数的52%。其中现实栖息地8片,占现实栖息地总数的80%,潜在栖息地16片,占潜在栖息地总数的44%。因此约半数的栖息地斑块和大部分现实栖息地斑块至少部分区域得到严格的保护。

|
| 图 1 四川山鹧鸪栖息地所涉及自然保护区分布示意图 Fig. 1 Sketch of nature reserve distribution in relation to Sichuan hill-partridges habitat |
|
美姑大风顶、马鞍山、麻咪泽、五莲峰这4个保护区分别将3个以上的四川山鹧鸪栖息地斑块部分地纳入保护管理范围,对于所涉及的栖息地斑块保护和斑块间栖息地重建修复可以起到积极作用。乌蒙山保护区保护了5片栖息地斑块的部分范围,但该保护区由3个独立片区构成,只有西部和中部2片包含了部分栖息地,保护作用受到限制。马边大风顶保护区虽然只涉及2个栖息地片区,但它保护了四川境内最大2片现实栖息地之间起关键连接作用的"大风顶北段"片(QZ11)潜在栖息地斑块东部,其重要意义值得一提。黑竹沟、老君山、芹菜坪、八月林、二十四冈、铜锣坝、白老林7个保护区只涉及单个栖息地斑块,但它们保护的是现实栖息地,尤其是老君山和芹菜坪是目前仅有的以四川山鹧鸪等雉类及其栖息地为主要保护对象的自然保护区,它们的建立对于该物种的保护管理起着重要作用。申果庄、老黎山、袁家湾、以那4个保护区处于分布区边缘,保护了部分潜在栖息地,有一定的辅助作用。
从具体的栖息地斑块来看,"甘洛-金口河-峨边-马边-美姑"片(XS01)和"绥江-永善-水富-盐津-大关"片(XS10)中各有4个保护区,"雷波-马边-屏山"片(XS03)有2个,其余现实栖息地斑块除2个无保护区外均各涉及1个保护区,可见3个最大的现实栖息地斑块得到较多保护区的支撑,有利于保护管理工作的开展。潜在栖息地中只有"甘洛南a"(QZ07)、"甘洛南b"(QZ08)和"大风顶北段"(QZ11)各得到2个保护区的保护,另有13片各涉及到1个保护区,其余20片没有保护区,可见潜在栖息地的保护力度不够。 3 讨论
四川山鹧鸪栖息地景观破碎化严重,亟待加以保护。首要是保护现有景观,防止进一步破碎化,然后在此基础上进行栖息地重建和扩增,消减栖息地构成单元之间的隔离因素以恢复和加强连通性。我们建议从以下几方面着手。
景观的保护应贯彻"预防为主"的原则,做好景观资源保护规划(毛文永,2001)。从栖息地斑块面积大小、形状完好程度、区位特殊性等方面确立保护优先级,按照各栖息地斑块的特征提出针对性的保护措施,再根据难易程度、投入大小、预期目标和效果等排出实施的时间安排,形成诸如保护管理计划一类的指导性文本,提交野生动物保护管理的相关政府部门及非政府组织,作为决策依据。如涉及栖息地的城乡建设规划、大型工程建设等应充分考虑该物种的保护,严格实行景观影响评估。
自然保护区在野生动植物保护,包括四川山鹧鸪及其同域分布物种的有效保护管理中起着重要的作用(Dai et al.,2009)。四川山鹧鸪分布区现有的17个保护区虽然初步形成了保护网络,但也只是保护了部分栖息地,保护区外还存在大面积栖息地需要加强保护。而且现有保护区的空间布局与该物种的分布格局存在差异,加之主要保护对象大多都不是四川山鹧鸪及其栖息地,区划的范围只能保护栖息地斑块的部分边缘地带,这种情况在凉山山系的几个保护区尤其明显。因此,今后可考虑调整主要保护对象包含本目标种,扩建现有保护区,将更多的四川山鹧鸪栖息地和适宜生境置于严格保护之下。为加强核心分布地的保护,时机成熟时应该在"甘洛-金口河-峨边-马边-美姑"片(XS01)现实栖息地所在的峨边、马边两县交界的大竹堡-洋溪河流域,以及"雷波-马边-屏山"片(XS03)现实栖息地所在的雷波县西宁河流域新建保护区(图 1)。此外还要做好保护区建立后的基础设施和人员能力建设工作。
对于边缘和孤立的栖息地斑块,在注意自身完整性的同时,如果周边附近存在有潜在连通性的其他斑块,可考虑建立走廊带,通过林地植被自然演替和人工促进更新恢复方式建立起实际的交流通道。特别应关注邻域分析中所述那些间隔很小的邻近斑块,公路交通可能不会完全阻隔斑块间的迁移交流,沿线的植被恢复、下穿隧道的开通等措施有利于廊道的维持和扩展,公路两侧的栖息地可望重新合一。
开展栖息地景观特征的动态监测,运用多时相遥感数据,结合相应的地面调查,定期了解栖息地变化情况。继续深入研究四川山鹧鸪生境需求、种群结构和扩散机制。此外运用景观遗传学方法研究破碎化栖息地景观特征与该物种种群格局和遗传多样性之间的关系,即种群遗传不连续性与景观特征不连续性的关联,获取景观特征与基因流、遗传漂变和选择等微进化过程之间相互作用的信息(薛亚东等,2011)。这些研究对四川山鹧鸪栖息地保护管理策略的制订、调整和具体措施的有效实施是必不可少的信息支撑。
| 戴波, 陈本平, 岳碧松, 等. 2014. 四川山鹧鸪栖息地分析与预测[J]. 四川动物, 33(3): 329-336. |
| 何芬奇, 沈尤, 张铭. 2008. 对四川山鹧鸪模式产地的再认识[J]. 动物分类学报, 33(3): 642-643. |
| 何芬奇, 宋道军, 金鑫, 等. 2009. 再谈四川山鹧鸪的模式产地[J]. 动物分类学报, 34(4): 946-948. |
| 李操, 余志伟, 胡杰. 2003. 四川山鹧鸪的分布和生境选择[J]. 动物学杂志, 38(6): 46-51. |
| 廖文波, 胡锦矗, 李操, 等. 2007. 繁殖期四川山鹧鸪鸣声行为特征[J]. 动物学研究, 28(1): 55-62. |
| 廖文波, 胡锦矗. 2010. 四川山鹧鸪生态习性研究进展[J]. 绵阳师范学院学报, 29(2): 67-71, 75. |
| 陆钢, 戴波, 李仁贵, 等. 2007. 四川老君山自然保护区的鸡形目种群密度研究[J]. 四川动物, 26(3): 572-576. |
| 毛文永. 2001. 景观资源的评价与保护[J]. 环境科学研究, 14(6): 54-56. |
| 薛亚东, 李丽, 吴巩胜, 等. 2011. 景观遗传学: 概念与方法[J]. 生态学报, 31(6): 1756-1762. |
| Dai B, Dowell SD, Garson P, et al. 2009. Habitat utilization by the threatened Sichuan partridge Arborophila rufipectus: consequences for managing newly protected areas in southern China[J]. Bird Conservation International, 19(2): 187-198. |
| Dai B, Dowell SD, Martins RP, et al. 1998. Conservation status of the Sichuan hill-partridge Arborophila rufipectus in China[J]. Bird Conservation International, 8(4): 349-359. |
| He LW, Dai B, Zeng B, et al. 2009. The complete mitochondrial genome of the Sichuan hill partridge (Arborophila rufipectus) and a phylogenetic analysis with related species[J]. Gene, 435(1-2): 23-28. |
| IUCN. 2014. The IUCN red list of threatened species[DB/OL]. (2014-2) [2014-5-24]. http://www.iucnredlist.org. |
| Liao WB, Fuller RA, Hu JC, et al. 2008a. Habitat use by endangered Sichuan partridges Arborophila rufipectus during the breeding season[J]. Acta Ornithologica, 43: 179-184. |
| Liao WB, Li C, Hu JC, et al. 2007. Habitat utilization of the Sichuan hill-partridge Arborophila rufipectus in the non-breeding season in Laojunshan Nature Reserve[J]. Zoological Research, 28(2): 172-178. |
| Liao WB, Li C, Hu JC, et al. 2008b. Roosting behaviour of the endangered Sichuan hill-partridge Arborophila rufipectus during the breeding season[J]. Bird Conservation International, 18(3): 260-266. |
| McGarigal K, Marks BJ. 1994. Fragstats. Spatial pattern analysis program for quantifying landscape structure. Version 2.0[CP/DK]. Corvallis: Forest Science Department, Oregon State University. |
| Rempel RS, Kaukinen D, Carr AP. 2012. Patch analyst and patch Grid[CP/OL]. Ontario Ministry of Natural Resources. Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario. http://www.cnfer.on.ca/SEP/patchanalyst/Patch5_1_Install.htm. |