 2015, Vol. 34(2)
2015, Vol. 34(2)扩展功能
文章信息
- 陈超, 杨志松, 潘载扬, 毕温磊, 陈鹏, Jacob R Owens, 侯蓉, 张志和, 古晓东, 齐敦武
- CHEN Chao, YANG Zhisong, PAN Zaiyang, BI Wenlei, CHEN Peng, Jacob R Owens, HOU Rong, ZHANG Zhihe, GU Xiaodong, QI Dunwu
- 圈养大熊猫发情期间行为变化和活动节律研究
- Behavioral Changes and Activity Rhythms of Captive Giant Pandas during the Breeding Season
- 四川动物, 2015, 34(2): 161-168
- Sichuan Journal of Zoology, 2015, 34(2): 161-168
- 10.3969/j.issn.1000-7083.2015.02.001
-
文章历史
- 收稿日期:2014-06-16
- 接受日期:2014-10-17






2. 成都大熊猫繁育研究基地,四川省濒危野生动物保护 生物学省部共建实验室, 成都 610086;
3. 四川省野生动物资源调查保护管理站, 成都 610000
2. Chengdu Research Base of Giant Panda Breeding, Sichuan Key Laboratory of Conservation Biology for Endangered Wildlife, Chengdu 610086, China;
3. Sichuan Management Station of Wildlife Resources Investigation and Protection, Chengdu 610000, China
动物行为是指在一定的环境条件下,动物为完成摄食排遗、体温调节、生存繁殖,以及满足其他生理需求而以一定姿势完成的一系列动作(蒋志刚等,2001),是对环境因素的影响和机体自身生理调节所做出的综合反应。同时,动物也可以根据周围环境条件(如光照、温度、湿度等)的变化及自身的生理状况来调整行为(Flannigan & Stookey,2002;Pépin et al.,2006;Piccione & Giannetto,2011),从而形成特定条件下的时间分配及活动节律模式(马飞雁等,2013)。昼夜活动节律是一种复杂的生物学现象,是动物对各种条件变化的一种周期性适应,既包括光、温度、湿度等非生物条件,也包括食物、种内社群关系和天敌等生物条件(孙儒泳,2001)。因此,研究珍稀濒危动物活动节律,既可丰富其生态学基础资料,又可为其保护与管理提供参考依据(马建章等,1999)。
发情期是动物最重要的生命时期之一,其行为特征和其他生命时期有差异(Verbeek,1972)。每种动物在发情期都有其最适的时间分配,且相对较短,因此,该时期相应的各种行为时间分配将对其种群繁衍产生重要影响(孙儒泳,2001)。大熊猫Ailuropoda melanoleuca发情期多在3月下旬至5月,雄性发情期可持续30 d左右,而雌性仅为1~3 d,且与雄性有不同步发情的现象(张志和,魏辅文,2006)。尽管目前在大熊猫繁育技术方面已取得较大突破,其圈养种群数量已达到了376只,然而,仍有超过1/3的个体不能自然交配。这些个体均属性不活跃个体,可能与雌雄发情是否同步、自身的生殖激素水平、食物营养条件、环境特征,以及社会状况等多种因素有关,而这些都与动物在发情期的时间分配和行为节律有紧密的联系(张志和,魏辅文,2006)。
动物昼夜活动时间分配对其繁殖有着深远影响(Daan & Aschoff,1982)。了解和掌握大熊猫的行为模式是对其进行科学研究和保护的重要前提(Long et al.,2004)。野外研究发现大熊猫昼夜有2个活动高峰,活动率约60%(胡锦矗,1987),且雌雄个体的活动节律有明显差异(Mainka & Zhang,1994),而这种节律受太阳辐射、温度等因子的影响(张晋东等,2011)。圈养条件下,大熊猫活动率则较野外有明显下降(约463%),存在3个活动高峰期和2个低谷(张志和,魏辅文,2006),表现为蹭阴、举尾、嗅阴和咩叫等行为明显增加(王雄清,1990),以及激素的变化,但受到其能量平衡的限制(聂永刚,2012)。
当前,大熊猫配对时间多是依据激素水平把握(Tian et al.,2007;于小杰等,2011),辅以发情行为来判断(杨胜林等,2007)。然而,激素测定结果往往具有一定的滞后性(Wasser,1996;刘娟等,2005),从而有可能错过最佳配对时间。同时,发情期行为的判断也依据饲养管理人员的主观经验,可能造成较大的主观误差。此外,对于大熊猫发情期的行为活动节律与时间分配尚缺乏系统的研究,而扫描取样法可以较好地评估行为发生节律和比例分配(兰天明等,2012)。为此,本研究采用全事件记录法和瞬时扫描取样法,以发情期圈养大熊猫为研究对象,对大熊猫的活动节律和行为变化进行研究,以期为圈养大熊猫自然交配率的提高提供行为学方面的理论依据和技术支持。 1 材料与方法 1.1 研究对象和地点
本实验以成都大熊猫繁育研究基地(以下简称熊猫基地)的月亮产房中8只成年大熊猫为研究对象,其中雄性2只(编号为M1和M2),雌性6只(分别编号为F1、F2、F3、F4、F5和F6)。M1和M2均为9岁,F1、F2、F3、F4、F5和F6分别为5岁、6岁、6岁、12岁、13岁和19岁,所有个体身体健康,且都有发情行为。截至2013年4月下旬,所有研究对象均完成交配或授精,其中M1、F1、F3、F4、F5和F6为自然交配,M2进行了人工采精,F2进行了人工授精。基于研究对象是否有交配史,把雌性个体分为初次发情(F1、F2、F3)和非初次发情(F4、F5、F6)。
大熊猫兽舍可分为室内活动场和户外活动场两部分,室内设有木床,户外设有乔木、灌木、草坪、栖架平衡木、水池及假山等丰容设施。每天09∶00将大熊猫放出户外,17∶30将其收进室内,喂食时间为09∶00和15∶30,竹子供应充足,期间间隔性地投食窝窝头、苹果、蜂蜜水和药。 1.2 研究方法 1.2.1 仪器和设备
在实验观察期间,用海康威视红外可调焦防水ICR日夜型筒型网络摄像机进行24 h观察。产房配种间内有固定摄像头(DS2CD8233F)、可移动摄像头(DS2CD8233F)、SONY摄像机(HDRPJ30E),数量分别为10台、10台和1台,并配有1台台式电脑(联想启天M7360)。配种间为长方形,在其上方4角以对角线形式安置固定摄像头以达到无盲区观察。可移动摄像头则放置舍外辅以进行行为观测,尽可能地减少人为干扰。 1.2.2 行为的观察与定义
采用全事件记录法,首先进行了9 d(2013年2月20—28日)的预观察期,每天观察2个时段,分别为上午09∶00—11∶00和下午14∶00—17∶30,利用摄像机观察与记录其行为,制定观察方案、记录表和定义行为谱。然后,通过实验方案的改进,进行第二阶段的实验观察与记录,时间为2013年3月1日—4月30日,利用摄像机和摄像头进行24 h行为观察,每只观察对象每周进行2 d的观测记录,采用瞬时扫描取样法每10 min记录一次行为状态。
参照胡锦矗等(1985)、刘定震等(1998,2002)、Snyder等(2003)、张志和和魏辅文(2006)对圈养大熊猫行为的定义和研究结果,将大熊猫外部表现行为分为6个类型:
摄食(Feeding):包括食用竹子、竹笋、窝窝头、苹果、水、蜂蜜和药物等。
求适(Grooming):先天获得性行为,以舔、舔毛(掌)、擦痒、抓痒为主的行为。
休息(Resting):个体以各种姿势保持静止状态,眼闭合或不闭合。
运动(Moving):包括刻板、走动、跑动、攀爬、游戏等行为。
探究(Exploring):盯住并缓慢地走到某一物体前,嗅闻或张望,有时在兽舍内会爬栏张望。
发情(Breeding):包括个体在发情期的蹭阴、露阴(针对雄性)、粪尿标记、戏水、举尾、脊柱前凸和交配行为。 1.2.3 数据处理
首先按照个体进行各个时间某种类型行为(如摄食、求适、休息、运动、探究和发情)取样数据的加和汇总;将每次扫描取样视为1个独立样本,以某个体的某行为发生次数与扫描取样中该个体总行为发生次数的比值作为该种行为在这一取样样本中所占的时间比例;然后将每天内扫描样本的数据平均,计算出每天的活动时间分配;最后用每天的平均值来表示该时期的活动时间分配。日活动节律则用主要行为类型在各个时间段(2 h)的频率平均值来表示。
其次,利用KolmogorovSmirnov检验对所有数据进行正态性检验,若数据符合正态分布,则采用单因素方差分析(OneWay ANOVA),反之,则采用MannWhitney U 检验,比较活动时间分配在个体间、不同性别间及初次发情与非初次发情间的差异,并依据数据分布特征选取Pearson相关性检验(正态)或Spearman相关性检验(非正态)进行不同行为类型间的相关性分析。数据分析在SPSS 180(杜荣骞,2009;陈胜可,2013)中完成,数值用Mean±SE表示。 2 结果 2.1 时间分配
在大熊猫的昼夜行为中,雄性最主要的行为是休息(4558%±225%,n=67),其余依次为摄食(2575%±175%,n=67)、运动(1858%±151%,n=67)、探究(435%±060%,n=67)、发情(311%±054%,n=67)和求适(263%±040%,n=67)(图 1)。和雄性相比,雌性休息行为更多(5319%±175%,ANOVA:F=7088,P<001);而运动(UTest:Z=-4280,P<001)、探究(UTest:Z=-2038,P<005)、求适(UTest:Z=-2928,P<001)和发情(UTest:Z=-4233,P<001)行为则明显减少,分别为1161%±097%、291%±032%、231%±071%和106%±017%;在摄食行为上则无差异(ANOVA:F=1518,P>005),为2892%±165%。

|
|
图 1 大熊猫发情期行为类型的昼夜时间分配
Fig. 1 Time budgets of giant p and as during the breeding season
“**”表示雌雄在该行为上的时间分配差异有高度统计学意义(P<001),“*”表示雌雄在该行为上的时间分配差异有统计学意义(P<005)。 Double asterisks indicate behavioral time budget between female and male was significantly different at P<001; one asterisk indicates behavioral time budget between female and male was significantly different at P<005. |
针对不同类型的雌性大熊猫(即初次发情和非初次发情),初次发情个体主要的行为是休息(5374%±238%,n=57),其余依次为摄食(2846%±224%,n=57)、运动(997%±111%,n=57)、求适(362%±142%,n=57)、探究(306%±045%,n=57)和发情(115%±025%,n=57);而与初次发情个体相比,非初次发情个体在休息(n=55)、摄食(n=55)、运动(n=55)、探究(n=55)、求适(n=55)和发情(n=55)行为上差异均无统计学意义(P>005),分别为5266%±259%(F=0093,df=1),2937%±242%(F=0076,df=1),1319%±155%(Z=-0915,df=1),277%±045%(Z=-0518,df=1),104%±020%(Z=-0865,df=1),097%±024%(Z=-0639,df=1)(图 2)。

|
| 图 2 雌性大熊猫发情期行为类型的昼夜时间分配 Fig. 2 budgets of female giant p and as during the breeding season |
雄性个体在发情期的运动行为出现了2个时间段(分别为08∶00—11∶00和15∶00—19∶00)的高峰期,而在13∶00、21∶00和05∶00为3个运动低谷期。总体来看,发情行为所占比例较少,主要发生在09∶00—12∶00和00∶00—02∶00的2个时段(图 3)。此外,摄食行为有3个高峰期,即09∶00—12∶00、15∶00—17∶00和23∶00—03∶00,而在12∶00—16∶00和20∶00—07∶00则表现为休息行为(图 3)。根据相关性检验分析,发现休息与摄食、求适、探究、运动和发情行为均呈极显著负相关(P<001,表 1);探究与运动行为显著正相关(P<001,表 1);发情与休息行为极显著负相关(P<001),而与运动(P<001)和探究行为(P<005)为显著正相关(表 1)。

|
| 图 3 雄性大熊猫发情期昼夜活动节律 Fig. 3 Activity rhythm of male giant p and as during the breeding season |
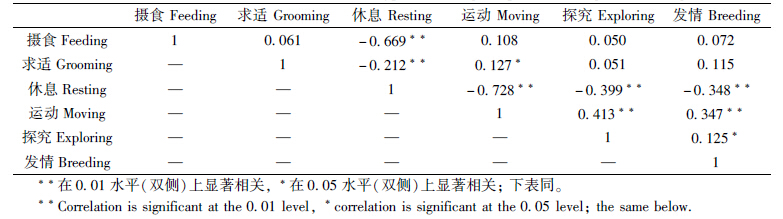
|
相比雄性大熊猫,初次发情和非初次发情雌性个体的运动行为均具有略晚的、相似的高峰期,分别为09∶00—11∶00和17∶00—19∶00,2活动低谷则具有一致性。摄食行为均有3个高峰期:09∶00—12∶00、17∶00—19∶00和23∶00—01∶00;发情行为发生在摄食行为之后,且时间较短,主要发生在07∶00—11∶00、15∶00—17∶00和01∶00—05∶00(图 4,图 5)。与雄性大熊猫相比,发情行为发生高峰及低谷期相似,但发生频率偏低(雄性最高频率为432%,而雌性为205%,图 6)。初次发情个体的休息行为与摄食、求适行为呈极显著负相关(P<001),运动行为与摄食行为呈极显著负相关(P<001);而发情行为与运动行为呈显著正相关(P<005,表 2)。非初次发情个体的休息行为与摄食、运动行为呈极显著负相关(P<001),而运动行为与探究行为极显著正相关(P<001),发情行为与摄食、求适行为显著正相关(P<005,表 3)。此外,具有发情交配经验个体的发情行为高峰期稳定,分别为09∶00、17∶00和01∶00;而初次发情个体则不稳定,发生高峰为09∶00—11∶00和15∶00—17∶00(图 7)。

|
| 图 4 初次发情雌性大熊猫发情期昼夜活动节律 Fig. 4 Activity rhythm of female giant p and as during the breeding season |

|
| 图 5 非初次发情雌性大熊猫发情期昼夜活动节律 Fig. 5 Activity rhythm of nonfirst oestrus female giant p and as during the breeding season |

|
| 图 6 雌雄大熊猫发情行为 Fig. 6 Oestrus behavior of giant p and as during the breeding season |
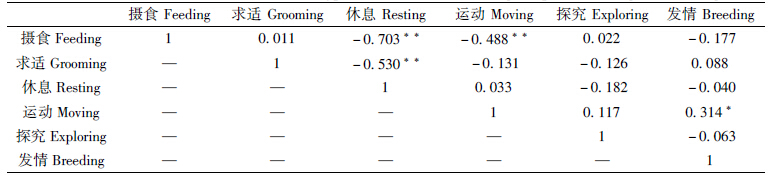
|
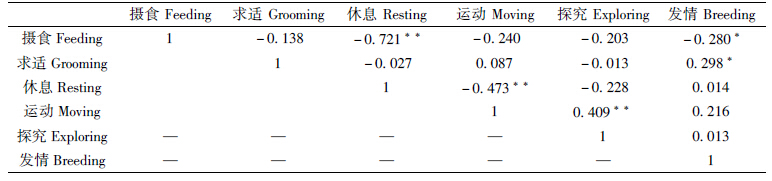
|

|
| 图 7 初次非初次发情雌性大熊猫发情行为 Fig. 7 Oestrus behavior of female giant p and as during the breeding season |
动物活动的时间分配与其代谢、能量利用有着密切关系,反映了长期进化形成的生存策略(孙儒泳,2001),从而形成了特定生理条件下的时间分配规律(Flannigan & Stookey,2002)。大熊猫主食竹类,且消化率很低,形成了能量简约型的活动时间分配和利用方式(Wei et al.,2000;胡锦矗,2001)。野生大熊猫的活动主要表现为觅食和游荡等,并决定其昼夜活动率,如春季成年雌性昼夜活动率为64%~65%,雄性则为69%(胡锦矗等,1985;胡锦矗,1987)。相对于野生大熊猫,圈养大熊猫的食物由饲养员定时供给,不需要自己觅食,休息和摄食行为成为其具有绝对优势的行为,有超过70%的时间用于这2种行为(张志和,魏辅文,2006)。多数其他动物繁殖期也会呈现这种时间分配规律(如草食性有蹄类,Brivio et al.,2010)。圈养大熊猫的活动时间较少(发情期雄性1858%,雌性997%),多数时间是在兽舍内或活动场游荡或刻板踱步。然而,由于体型和繁殖角色的不同,大熊猫两性在能量利用和栖息地利用方面也有不同(Qi et al.,2011),雌性和雄性均以休息为主,然而,雌性较雄性在休息行为上有明显增加,发情、运动、探究和求适行为则明显减少。雌性发情期间的运动行为高峰期略晚(Mainka & Zhang,1994),这可能与其性二型的节省能量策略有关。当然,随着对圈养条件的适应,其活动节律与野生大熊猫也有一定的差异(胡锦矗,2001)。
动物活动的时间分配是其对环境条件的一种适应,受遗传、食物能量、性别和繁殖状况等因素影响(孙儒泳,2001)。在圈养条件下,由于空间及资源有限,动物的生理需求得不到满足,从而导致其多种行为都表现出明显的人为影响的特征(Mainka & Zhang,1994)。比如,在上午(09∶00—11∶00)和下午(15∶00—17∶00),圈养大熊猫摄食和运动规律与食物的投放时间(09∶00和15∶30)具有高度的相关性。对圈养环境形成的这种适应性行为随着圈养时间的延长亦难以消失(张志和,魏辅文,2006)。
动物在发情期的发情行为表现明显,其特点与动物体的能量平衡有关(孙儒泳,2001)。在发情期,大熊猫与其发情相关的活动明显增加,休息行为有所减少(胡锦矗,2001)。然而,大熊猫发情行为所占时间比例较低(图 1),主要集中在上午、下午及凌晨3个高峰期。大熊猫在发情期的发情行为与活动量增加的行为(如运动和探究行为)呈正相关,但仅雄性大熊猫发情行为与休息行为有极显著负相关性,雌性则无显著相关性。同样,发情期的雄性小熊猫Ailurus fulgens较雌性更活跃,活动频率更高(刘雪卿等,2004)。对于雌性大熊猫不同的繁殖状况来说,其活动时间分配并未发现明显差异,非初次发情个体仅表现在摄食和运动行为略有增加,这或许与其不同年龄间能量利用效率差异有关(刘定震等,2002;张志和,魏辅文,2006)。
当前,尽管圈养大熊猫的规模已达376只,但仍然面临着诸多繁育方面的难题,如圈养种群质量有待提高、遗传多样性较低、自然交配率较低和种公兽缺乏等问题(张志和,魏辅文,2006)。了解大熊猫在发情期昼夜行为的变化规律和时间分配特点,可更好地掌握种公兽的行为模式,确定人工授精时间,从而提高自然交配的繁殖成功率。
致谢:承蒙成都大熊猫繁育研究基地黄祥明、吴孔菊、邓陶、张浩、程瑛等的支持和帮助,在此一并表示感谢!
| 陈胜可. 2013. SPSS统计分析从入门到精通(第二版)[M]. 北京: 清华大学出版社. |
| 杜荣骞. 2009. 生物统计学(第三版)[M]. 北京: 高等教育出版社. |
| 胡锦矗, Schaller GB, 潘文石, 等. 1985. 卧龙的大熊猫[M]. 成都: 四川科学技术出版社. |
| 胡锦矗. 1987. 大熊猫的昼夜活动规律[J]. 兽类学报, 7(4): 241-245. |
| 胡锦矗. 2001. 大熊猫研究[M]. 上海: 上海教育科技出版社. |
| 蒋志刚, 李春旺, 彭建军, 等. 2001. 行为的结构、刚性和多样性[J]. 生物多样性, 9(3): 265-274. |
| 兰天明, 田银平, 胡晓楠, 等. 2012. 目标取样法和扫描取样法在圈养黑熊行为研究中的比较[J]. 经济动物学报, 16(2): 94-101. |
| 刘定震, 房继明, 孙儒泳, 等. 1998. 大熊猫个体不同性活跃能力的行为比较[J]. 动物学报, 44(1): 27-34. |
| 刘定震, 张贵权, 魏荣平, 等. 2002. 性别与年龄对圈养大熊猫行为的影响[J]. 动物学报, 48(5): 585-590. |
| 刘娟, 陈玥, 郭丽然, 等. 2005. 圈养大熊猫刻板行为观察及其激素水平测定[J]. 北京师范大学学报: 自然科学版, 41(1): 75-78. |
| 刘雪卿, 张泽钧, 魏辅文, 等. 2004. 圈养小熊猫繁殖行为变化及繁殖行为对策[J]. 兽类学报, 24(2): 173-176. |
| 马飞雁, 余晓俊, 陈珉, 等. 2013. 圈养獐春夏季昼间行为时间分配及活动节律[J]. 兽类学报, 33(1): 28-34. |
| 马建章, 徐利, 张洪海, 等. 1999. 大兴安岭地区紫貂的活动节律[J]. 兽类学报, 19(2): 95-100. |
| 聂永刚. 2012. 秦岭野生大熊猫繁殖生态学研究[D]. 北京: 中国科学院动物研究所. |
| 孙儒泳. 2001. 动物生态学原理(第三版)[M]. 北京: 北京师范大学出版社. |
| 王雄清. 1990. 大熊猫繁殖研究进展[J]. 四川师范学院学报: 自然科学版, 11(2): 118-123. |
| 杨胜林, 李敏, 周强, 等. 2007. 雌性大熊猫发情行为观察与相关指标分析[J]. 四川动物, 26(3): 664-668. |
| 于小杰, 胡德夫, 唐勇清, 等. 2011. 大熊猫粪便类固醇激素研究进展[J]. 生物学通报, 46(5): 1-2. |
| 张晋东, Hull V, 黄金燕, 等. 2011. 大熊猫取食竹笋期间的昼夜活动节律和强度[J]. 生态学报, 31(10): 2655-2661. |
| 张志和, 魏辅文. 2006. 大熊猫迁地保护理论与实践[M]. 北京: 科学出版社. |
| Brivio F, Grignolio S, Apollonio M. 2010. To feed or not to feed? Testing different hypotheses on rut-induced hypophagia in a mountain ungulate[J]. Ethology, 116(5): 406-415. |
| Daan S, Aschoff J. 1982. Circadian contributions to survival[M].// Aschoff J, Daan S, Groos GA(eds). Vertebrate circadian systems: structure and physiology. Berlin: Springer-Verlag: 305-321. |
| Flannigan G, Stookey JM. 2002. Day-time time budgets of pregnant mares housed in tie stalls: a comparison of draft versus light mares[J]. Applied Animal Behaviour Science, 78(2): 125-143. |
| Long Y, Lü Z, Wang DJ, et al. 2004. Nutritional strategy of giant pandas in the Qinling mountains of China[M].// Lindburg DG, Baragona K (eds). Giant pandas: biology and conservation. Berkeley, California: University of California Press: 90-100. |
| Mainka AS, Zhang HM. 1994. Daily activity of captive giant pandas (Ailuropoda melanoleuca) at the Wolong Reserve[J]. Zoo Biology, 13(1): 13-20. |
| Pépin D, Renaud PC, Dumont B, et al. 2006. Time budget and 24-h temporal rest-activity patterns of captive red deer hinds[J]. Applied Animal Behaviour Science, 101(3): 339-354. |
| Piccione G, Giannetto C. 2011. State of the art on daily rhythms of physiology and behaviour in horses[J]. Biological Rhythm Research, 42(1): 67-88. |
| Qi DW, Zhang SN, Zhang ZJ, et al. 2011. Different habitat preferences of male and female giant pandas[J]. Journal of Zoology (London), 285(3): 205-214. |
| Snyder RJ, Zhang AJ, Zhang ZH, et al. 2003. Behavioral and developmental consequences of early rearing experience for captive giant pandas (Ailuropoda melanoleuca)[J]. Journal of Comparative Psychology, 117(3): 235-245. |
| Tian H, Wei RP, Zhang GQ, et al. 2007. Age differences in behavioral responses of male giant pandas to chemosensory stimulation[J]. Zoological Research, 28(2): 134-140. |
| Verbeek NAM. 1972. Daily and annual time budget of the yellow-billed magpie[J]. The Auk, 89(3): 576-582. |
| Wasser SK. 1996. Reproductive control in wild baboons measured by fecal steroids[J]. Biology of Reproduction, 55(2): 393-399. |
| Wei FW, Feng ZJ, Wang ZW, et al. 2000. Habitat use and separation between the giant panda and the red panda[J]. Journal of Mammalogy, 81(2): 448-455. |