 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 田华香, 胡凤霞, 胡海洋, 燕帅国, 赵晓进
- TIAN Huaxiang, HU Fengxia, HU Haiyang, YAN Shuaiguo, ZHAO Xiaojin
- 太行山猕猴颅骨变量的波动不对称性
- Fluctuating Asymmetry of Skull Characters of Macaca mulatta in Taihang Mountains
- 四川动物, 2015, 34(4): 500-504
- Sichuan Journal of Zoology, 2015, 34(4): 500-504
- 10.11984/j.issn.1000-7083.2015.04.004
-
文章历史
- 收稿日期:2014-10-31;接受日期:2015-02-25






2. 河南师范大学生命科学学院, 河南新乡 453007
2. College of Life Sciences, Henan Normal University, Xinxiang, Henan Province 453007, China
Mather(1953)提出波动性不对称(fluctuating asymmetry,FA),是指生命体具有对称性结构的性状在发育过程中出现了细微的随机偏离,导致对称结构出现了某种程度的不对称现象。FA是生命科学研究中常用的专业术语之一。目前认为FA主要是生物在发育过程中受到环境压力、发育稳态及遗传因素共同作用的结果,其遗传力极低(Manning & Ockenden,1994)。很明显如果某些生物个体的对称性状越精确,说明这些个体能够对不断变化的内外环境做出更精细的调节,发育状况良好,具有较强的抗干扰能力(van Valen,1962)。FA的水平可用来指示某一种群对环境压力的进化和适应能力(杨旭等,2009)。FA可作为个体发育稳定性的一个标记。Hallgrimsson(1993)观察测量138例食蟹猴6个颅骨对称变量的FA,结果表明颅骨变量的FA与年龄有关。
王凤产等(2012)报道了太行山猕猴Macaca mulatta掌骨和跖骨的FA,指出掌骨和跖骨骨干截面积的FA值相对较大,长度变量的FA值较小;FA值存在一定程度的性差;推测猕猴掌骨和跖骨FA特征是种群水平上的。程如意等(2012)报道了太行山猕猴四肢长骨的不对称性,结果表明肱骨存在方向性不对称,显示出左侧优势的趋势,第二掌骨显示右偏趋势,股骨作为对照组不对称性较弱。有关太行山猕猴颅骨形态特征已有许多资料报道(赵晓进等,1997,1999a,1999b,2001,2003,2004),这些研究主要集中在性差、异速生长和体征等方面。目前国内尚未见到有关猕猴颅骨FA的报道。本文对太行山猕猴颅骨FA进行观察研究,以期为太行山猕猴骨骼不对称研究提供基本数据资料。 1 材料和方法
猕猴颅骨标本62例(♀43,♂19),干燥完整,现存于河南师范大学生物标本馆,将标本按性别和年龄分组(Schultz,1958)。性别组:雌43例,雄19例。年龄组:亚成体组19例(齿式为2121或2122),成体组43例(齿式为2123)。
变量选择见Schultz(1958)和Hallgrimsson(1993)的报道。用电子数字游标卡尺对两侧颅骨的24项变量进行测量,单位为mm,精度为0.02 mm。本课题组对太行山猕猴颅骨形态学已经做了一些相关研究,掌握了骨骼测量的基本技术并积累了许多经验。所有变量都由经验丰富和经过培训的专业技术人员进行测量。另外,在同一环境的不同时间中进行重复测量,2次测量的相关系数均大于0.96。可以排除人为因素导致的测量误差。
为了消除颅骨变量绝对值大小可能对FA产生的影响,FA需要做标准化转换,计算公式(Hallgrimsson,1993;王凤产等,2012):
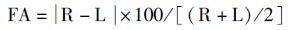
L为颅骨左侧某一变量,R为颅骨右侧相对应的变量。
数据采用SPSS 20.0软件进行处理。
采用Kolmogorov-Smirnov方法进行正态分布检验。由于大部分FA值不符合正态分布(P<0.05),所以颅骨变量的FA描述性统计采用中间值作为统计量,组间差异采用非参数Mann-Whitney U test检验。变量的字母标记和测量位点见图 1和表 1。

|
| 图 1 太行山猕猴颅骨变量测量位点(字母标识见表 1) Fig. 1 Measurement made on the skull of Macaca mulattain Taihang Mountains |
| 变量
Variable | 距离
Distance | 描述
Description | 注释
Note |
| X1 | a-z | Intradentale superior-zygion | 口点-颧骨点 |
| X2 | a-u | Intradentale superior-fronto malare | 口点-额颧缝 |
| X3 | b-q | Nasale-infraorbital foramen | 鼻突-眶下孔 |
| X4 | b-z | Nasale-zygion | 鼻突-颧骨点 |
| X5 | b-u | Nasale-fronto malare | 鼻突-额颧缝交点 |
| X6 | c-u | Nasion-fronto malare | 鼻根点-额颧缝交点 |
| X7 | d-t | Bregma-asterion | 前卤点-颞枕缝交点 |
| X8 | d-w | Bregma-porion | 前卤点-外耳道上缘点 |
| X9 | g-e | Suturae palatina mediana-the outer edge of the first molars | 腭正中缝-第一臼齿外缘 |
| X10 | g-f | Suturae palatina mediana-ectomalare | 腭正中缝-第二臼齿外缘 |
| X11 | g-y | Suturae palatine mediana-inferior zygo-temporal suture | 腭正中缝-颞颧缝交点 |
| X12 | h-y | Posterior nasal spine-inferior zygo-temporal suture | 鼻棘下点-颞颧缝交点 |
| X13 | k-w | Basion-porion | 颅底点-外耳道上缘点 |
| X14 | k-j | Basion-foramina spinosum | 颅底点-棘孔 |
| X15 | l-i | Opisthion-carotid canal | 枕外嵴点-颈动脉管 |
| X16 | l-w | Opisthion-porion | 枕外嵴点-外耳道上缘点 |
| X17 | m-p | Frontomalare orbitale-zygomaxillare superior | 眶额颧点-颧颌上点 |
| X18 | n-o | Dakryon-ektokonchion | 眶内缘点-眶外缘点 |
| X19 | r-w | Lambda-porion | 人字点-外耳道上缘点 |
| X20 | r-t | Lambda-asterion | 人字点-颞枕缝交点 |
| X21 | s-t | Inion-asterion | 枕外隆突-颞枕缝交点 |
| X22 | x-y | Anterior external auditoory meatus-inferior zygo-temporal suture | 外耳道前缘点-颞颧缝交点 |
| X23 | x-v | Anterior external auditoory meatus-pterion | 外耳道前缘点-翼点 |
| X24 | u-y | Fronto malare-inferior zygo-temporal suture | 额颧缝-颞颧缝交点 |
性别组颅骨FA值的描述性统计和非参数Mann-Whitney U test结果见表 2(左半部分)。大部分FA值在性别间的差异无统计学意义(P>0.05),X4和X22在性别间的差异有统计学意义(P<0.05)。此外,2个变量X12和X20在性别间的差异接近统计学水平,P值分别为0.061和0.083。总体上结果显示FA值是雌性大于雄性(18/24,75.1%)。4个变量(包括有统计学差异和接近统计学差异)中3个的FA值是雌性大于雄性。
| 变量
Variable | 雄性
Male(Median) | 雌性
Female(Median) | Z | P | 亚成体
Subadult(Median) | 成体
Adult(Median) | Z | P |
| X1 | 1.2869 | 1.4009 | -0.481 | 0.631 | 1.9509 | 1.2838 | -1.117 | 0.264 |
| X2 | 0.9044 | 1.1196 | -0.496 | 0.620 | 1.7340 | 0.9355 | -1.907 | 0.056 |
| X3 | 2.8106 | 2.4865 | -0.893 | 0.372 | 2.6625 | 2.7318 | -0.155 | 0.877 |
| X4 | 1.0011 | 1.6875 | -2.588 | 0.010* | 1.6475 | 1.3441 | -0.078 | 0.938 |
| X5 | 1.4723 | 1.7747 | -0.267 | 0.789 | 2.0008 | 1.4073 | -2.063 | 0.039* |
| X6 | 1.3333 | 1.9460 | -1.641 | 0.101 | 1.8475 | 1.5629 | -0.419 | 0.675 |
| X7 | 1.6581 | 2.2033 | -0.909 | 0.364 | 2.4943 | 1.7830 | -0.729 | 0.466 |
| X8 | 1.5061 | 1.7540 | -0.924 | 0.356 | 1.6240 | 1.7593 | -0.326 | 0.745 |
| X9 | 1.8174 | 1.9256 | -0.450 | 0.652 | 1.9547 | 1.8194 | -0.558 | 0.577 |
| X10 | 1.8080 | 1.9211 | -0.252 | 0.801 | 1.8451 | 1.8779 | -1.318 | 0.187 |
| X11 | 1.5029 | 1.4201 | -0.176 | 0.861 | 1.4771 | 1.4615 | -0.341 | 0.733 |
| X12 | 0.9210 | 1.3470 | -1.870 | 0.061 | 1.0052 | 1.2876 | -0.512 | 0.609 |
| X13 | 1.1981 | 1.3896 | -0.191 | 0.849 | 1.1387 | 1.5868 | -0.713 | 0.476 |
| X14 | 3.2677 | 1.6764 | -1.519 | 0.129 | 1.4694 | 2.6310 | -1.179 | 0.239 |
| X15 | 1.0900 | 1.9448 | -1.099 | 0.272 | 1.4265 | 1.7898 | -0.915 | 0.360 |
| X16 | 1.4856 | 1.4899 | -0.191 | 0.849 | 1.5646 | 1.4878 | -0.233 | 0.816 |
| X17 | 2.3269 | 2.0130 | -0.512 | 0.609 | 1.8547 | 2.2211 | -0.155 | 0.877 |
| X18 | 1.6012 | 2.0455 | -1.183 | 0.237 | 2.1645 | 1.6244 | -1.334 | 0.182 |
| X19 | 1.8561 | 1.8613 | -0.878 | 0.380 | 1.2979 | 1.8948 | -1.055 | 0.292 |
| X20 | 2.4887 | 3.3473 | -1.733 | 0.083 | 2.9477 | 3.2921 | -0.775 | 0.438 |
| X21 | 2.5182 | 2.7890 | -0.328 | 0.743 | 2.5240 | 2.9482 | -0.233 | 0.816 |
| X22 | 2.7084 | 1.4760 | -1.962 | 0.048* | 1.6790 | 2.0805 | -0.341 | 0.733 |
| X23 | 3.0512 | 2.5981 | -0.725 | 0.468 | 3.3104 | 2.4917 | -0.946 | 0.344 |
| X24 | 1.5462 | 1.9496 | -0.786 | 0.432 | 2.5503 | 1.6089 | -1.830 | 0.067 |
| 注: *P<0.05,Z值表示2个独立样本非参数检验的Mann-Whitney U test的检验统计量。 Notes: *P<0.05,Z score indicates test statistic of two independent samples of non-parametric test from Mann-Whitney U test. |
年龄组颅骨FA值的描述性统计和非参数Mann-Whitney U test结果见表 2(右半部分)。大部分FA值在年龄组间的差异无统计学意义(P>0.05),X5在年龄组间的差异有统计学意义(P<0.05),X2(P=0.056)和X24(P=0.067)在年龄间的差异接近统计学水平。总体上结果显示FA值在年龄组间的差异较小。3个变量的FA值(包括有统计学差异和接近统计学差异)都是亚成体组大于成体组。结果提示年龄组间FA值差异较小,有亚成体组大于成体组的趋势(13/24,54.4%)。 3 讨论
目前,FA研究已经成为生物学、人类学、医学和行为学所关注的热点。研究表明,人类的某些FA与某些疾病密切相关,例如,指长波动不对称与男性不育有关(陆宏等,2011,2012),精神分裂症患者的皮纹嵴线数FA水平明显高于正常人群(党洁等,2007)。研究猕猴颅骨波动性不对称的目的和意义在于:(1)目前,国内非人灵长类FA研究较少,颅骨FA研究更少,本研究可以填补国内有关猕猴颅骨FA研究资料的匮乏;(2)探讨猕猴颅骨变量FA模式和FA变化趋势,提供非人灵长类之间进行比对的数据;(3)探讨年龄、性别等因素对猕猴颅骨FA变化的影响。
许多专家学者坚持认为动物的FA可以作为群体水平上发育是否稳定或机体发育中抗各种环境因素干扰能力的一个重要指标(Knierim et al.,2007)。动物的饲养方式、自然环境、社会地位和化学污染等因素可以明显影响动物对称结构的发育稳定性(Klingenberg,2003)。FA是自然界普遍存在的现象,也是生命体的自然属性之一。
Stub等(2002)研究了3个小鼠品系在不同饲育环境下的FA变化,指出其中有2个品系由于环境的影响导致FA值明显降低。Manning和Ockenden(1994)报道了具有良好对称性的赛马具有更强的奔跑能力。本课题组的其他成员(程如意等,2012;王凤产等,2012)先后报道了猕猴长骨和掌跖骨方向不对称和FA的变化。
本研究结果显示,总体上颅骨FA值在性别组间和年龄组间的差异较小,大部分变量在组间比较没有统计学意义。少数颅骨变量性别组间有统计学差异或接近统计学差异,结果提示雌性的FA值有大于雄性的趋势。年龄组间比较显示也有少量变量有统计学差异或接近统计学差异,结果提示亚成体组的FA值可能大于或等于成体组。
Hallgrlmsson(1993)研究了食蟹猴颅骨的FA,结果显示性别组间没有差异。本研究结果与其相似。Seth(1969)对懒猴和蜂猴的锁骨和长骨的FA进行了研究,结果表明:几乎所有的长骨不对称值都是雌性大于雄性。但是他们没有对这一结果进行解释。Falk等(1988)观察了150例猕猴四肢长骨FA模式,结果也显示出长骨大部分变量的不对称值是雌性大于雄性。Helmkamp和Falk(1990)对137例猕猴骨骼FA进行研究,结果表明雌性尺骨的某些变量不对称性明显大于雄性(P=0.045)。并提出雄性骨骼不对称变化随着年龄的增长而减少,而雌性骨骼不对称变化是随着年龄增长而增加。因此推测雄性和雌性猕猴骨骼不对称性的发育可能存在不同的模式。Hallgrlmsson(1993)研究食蟹猴颅骨不对称时提出FA的增加与发育的时间并不是完全均衡的,这种差异主要反映了胚胎发育早期一些环境条件的变化。本研究结果支持这一模式。
另外作者分析成年雌性FA值较大,可能与其特殊生理活动(如月经周期、妊娠、分娩和哺乳等)和在群体中社会地位等有关。有资料表明太行山猕猴是一个等级分明、分工明确的社会群体,大部分雌性个体在群体中处于较低的社会地位(苏亚,2012)。
年龄组间部分变量的FA值有随着年龄的增加而减少的趋势。X5在年龄组间的差异有统计学意义(P<0.05)。亚成体组FA值明显大于成体组。此结果可能与变量的位置或变量的类型有关,也反映出整体上FA值从亚成体到成体过渡阶段的特征。有资料支持并解释了这一结果。Ferguson(1986)报道了一种杂合体鳟鱼身体发育不对称特征,指出这种鱼的FA和生长发育速率呈负相关。Hallgrlmsson(1993)对192例食蟹猴颅骨变量FA进行观察,提出了食蟹猴的生长发育对FA产生一定的影响。他认为哺乳动物两侧对称结构会出现轻微的偏差,主要是由于胚胎发育的环境不同和出生后机械刺激的不对称所致,它们对骨骼FA的影响大小取决于器官与环境之间的相互作用,很明显生长速度越快,FA值越大。所以在个体发育过程中,FA一般随着身体大小和相对生长速率增加而逐渐减弱。因此,这种发育干扰因素的特点是早期较小的个体具有较高的FA。然而,还有一些试验结果表明FA与发育时间呈正相关。证据来自Saunders和Mayhall(1982)报道人类牙齿形态不同发育阶段和FA呈正相关。Kobyliansky和Livshits(1989)报道人类某些骨骼形态的FA和年龄呈正相关。有关年龄或身体大小与FA的关系是比较复杂多变的,需要进一步在不同种群中去观察和验证。
另外,因为本研究的年龄组只有亚成体组和成体组,两组间的个体大小差别不是很大,分组的依据主要是根据牙齿萌发情况,可能导致组间的体型差异较小。这里选择较多的颅骨不对称变量主要是为了更好地了解颅骨FA的总体模式,因为影响颅骨FA的因素很多。
总之,在讨论非人灵长类骨骼包括颅骨不对称变化模式或规律时,必须考虑到可能存在的各种因素,例如:遗传和非遗传性因素、出生前和出生后的环境因素(如激素作用和机械负荷)、不同部位的骨骼(如颅骨,下颌骨和肱骨等)和同一骨骼的不同变量(股骨的长度和粗壮度)、个体生长发育速率和性别差异、人类和非人灵长类种属特异性等。毫无疑问,人们所观察到的不对称模式是上述各种影响因素交互作用的结果。因此,不能简单地推测FA主要与大脑结构不对称、利手行为和出生后肢骨接受两侧机械刺激不对称有关。
| 程如意, 王凤产, 胡晓林, 等. 2012. 猕猴长骨的不对称性研究[J]. 河南师范大学学报, 40(1): 130-132. |
| 党洁, 霍正浩, 彭亮, 等. 2007. 精神分裂症患者皮纹a-b嵴线数波动性不对称的研究性[J]. 人类学学报, 26(1): 64-68. |
| 陆宏, 刘永杰, 霍正浩, 等. 2011. 生物体波动性不对称与人类疾病的研究进展[J]. 宁夏医科大学学报, 33(9): 899-992. |
| 陆宏, 党洁, 霍正浩, 等. 2012. 指长波动性不对称与男性不育的相关性[J]. 人类学学报, 31(1): 82-87. |
| 苏亚. 2012. 太行山猕猴肢骨形态研究[D]. 北京: 中国地质大学: 34-44. |
| 王凤产, 胡晓林, 赵晓进, 等. 2012. 太行山猕猴掌骨和蹠骨的波动性不对称[J]. 江西师范大学学报, 36(5): 547-550. |
| 杨旭, 刘云波, 孙淑华, 等. 2009. 实验动物产生波动性不对称的影响因素[J]. 中国比较医学杂志, 19(1): 63-66. |
| 赵晓进, 张红绪, 李卫国, 等. 1997. 太行山的猕猴头骨和某些体征研究初报[J]. 动物学报, 43(增): 123-125. |
| 赵晓进, 翟心慧, 张新胜, 等. 1999a. 太行山猕猴颅骨变量的相关性研究[J]. 解剖学杂志, 22(2): 174-177. |
| 赵晓进, 李卫国, 张红绪, 等. 1999b. 成年太行山猕猴颅骨变量回归分析[J]. 河南师范大学学报, 27(1): 77-79. |
| 赵晓进, 罗艳蕊, 李荧, 等. 2001. 太行山猕猴颅骨变量主成分分析[J]. 河南师范大学学报, 29(2): 54-56. |
| 赵晓进, 雷梦云, 滑兵, 等. 2003. 不同灵长类颅骨变量主成分分析[J]. 湛江师范学院学报, 24(6): 55-59. |
| 赵晓进, 翟鹏飞, 张红绪, 等. 2004. 猕猴颅骨变量和面角的性差研究[J]. 河南师范大学学报, 32(4): 83-86. |
| Ferguson MM. 1986. Developmental stability of rainbow trout hybrids: genomic coadaptation or heterozygosity[J]. Evolution, 40(2): 323-330. |
| Falk D, Pyne L, Helmkamp RC, et al. 1988. Directional asymmetry in the forelimb of Macaca mulatta[J]. American Journal of Physical Anthropology, 77(1): 1-6. |
| Helmkamp RC, Falk D. 1990. Age-and sex-associated variations in the directional asymmetry of rhesus macaque forelimb bones[J]. American Journal of Physical Anthropology, 83(2): 211-218. |
| Hallgrlmsson B. 1993. Fluctuating asymmetry in Macaca fascicularis: a study of the etiology of developmental noise[J]. International Journal of Primatology, 14(3): 421-443. |
| Kobyliansky E, Livshits G. 1989. Age dependent changes in morphometric and biochemical traits[J]. Annals of Human Biology, 16(3): 237-247. |
| Klingenberg CP. 2003. Fluctuating asymmetry and animal welfare: how far are we? and how far should we go?[J]. The Veterinary Journal, 166(1): 5-6. |
| Knierim U, Van Dongen S, Forkman B, et al. 2007. Fluctuating asymmetry as an animal welfare indicator-a review of methodology and validity[J]. Physiology and Behavior, 92(3): 398-421. |
| Mather K. 1953. Genetical control of stability in development[J]. Heredity, 7(3): 297-299. |
| Manning JT, Ockenden L. 1994. Fluctuating asymmetry in racehorses[J]. Nature, 370(6486): 185-186. |
| Schultz AH. 1958. Craninal and dental varibility in Colobus monkeys[J]. Journal of Zoology, 130(1): 79-105. |
| Seth PK. 1969. Asymmetries of the long bones of the limbs and of the clavicles in Indian Lorises[J]. Primates, 10(2): 163-170. |
| Saunders SR, Mayhall JT. 1982. Fluctuating asymmetry of dental morphological traits: new interpretations[J]. Human Biology, 54(4): 789-799. |
| Stub C, Ritskes HM, Thon R, et al. 2002. Fluctuating asymmetry in mice and rats: evaluation of the method[J]. Laboratory Animals, 36(2): 193-199. |
| van Valen L. 1962. A study of fluctuating asymmetry[J]. Evolution, 16(2): 125-142. |