 2012, Vol. 70
2012, Vol. 70中国气象学会主办。
文章信息
- 段居琦, 周广胜. 2012.
- DUAN Juqi, ZHOU Guangsheng. 2012.
- 中国单季稻种植北界的初步研究
- A preliminary study of planting northern boundary of single harvest rice in China
- 气象学报, 70(5): 1165-1172
- Acta Meteorologica Sinica, 70(5): 1165-1172.
- http://dx.doi.org/10.11676/qxxb2012.098
-
文章历史
- 收稿日期:2011-07-11
- 改回日期:2012-03-05




2. 南京信息工程大学大气物理学院, 南京, 210044;
3. 中国科学院植物研究所植被与环境变化国家重点实验室, 北京, 100093
2. School of Atmospheric Physics, Nanjing University of Information Science and Technology, Nanjing 210044, China;
3. State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
中国是世界水稻栽培大国,也是稻作起源国之一。研究(Gong et al,2007)指出,长江中下游地区可能是中国稻作发源地,至史前为止,水稻由此分3波传向中国的其他地区及周边朝鲜、日本等国,其传递到最北方的位置是辽东半岛。如今,水稻已在中国广泛分布。黑龙江省是中国重要的粮食产区(程叶青等,2005),是目前单季稻种植最北部的省份,同时也是水稻生产对气候变化最为敏感的地区之一(朱晓禧等,2008)。
由于水稻种植在中国无南界,因此,水稻种植的北界和海拔高度成为水稻种植关注的焦点(云雅如等,2007)。科学地确定水稻种植北界,可为调整水稻生产布局、扩大水稻栽培区、充分利用气候资源和评估水稻种植区对气候变化的敏感性提供依据。目前,确定水稻种植临界区的方法主要可分为两类:一类是根据温度对水稻的限制作用,确定温度的临界条件(中国农业科学院,1986; 中国农林气候区划组,1987)。这类方法主要包括中国农业科学院确定的以2000℃·d积温作为单季稻分布的临界热量条件和中国农林气候区划协作组确定的以稳定通过10℃持续日数≥110 d和稳定通过18℃持续日数≥30 d作为单季稻种植的临界热量条件。另一类是根据实地调查与气候条件分析结合的方法,确定水稻种植临界区域(江爱良,1982; 张谊光,1983)。这类方法在确定中国水稻种植上界中有应用,是通过 实地调查和对水稻生长的气候生态条件进行分析,认为中国水稻种植的上界位于云南丽江的宁蒗县境内,稻田海拔达2660 m,其与尼泊尔的朱姆拉可能是世界上种植水稻最高的地区(江爱良,1982)。有研究(浙江农业大学,1981)推测,中国单季稻实际种植北界可达黑龙江漠河地区,超过了基于温度限制方法确定的单季稻种植北界,但这一结论没有得到证实。
熵反映了物质系统状态的复杂程度,最大熵原理的核心思想是基于有限的已知信息对未知分布进行无偏推断的一种数学方法(Jaynes,1957),已应用于气象学和生态学等领域(张学文等,1995; 邢丁亮等,2011)。在气象研究中,根据最大熵原理揭示气压和温度服从均匀分布,比湿、位能、风速服从负指数分布,总能量、位温服从Г分布(周家斌等,1997);根据最大熵原理推导的稳定层云云滴谱方程(张学文,1987a; 郑国光,1988),也用于降水概率分布、雨深的面分布和气压的面分布等方面(张学文,1987b)。在生态研究中,最大熵原理已广泛用于预测物种相对多度(Shipley,2010)、物种地理分布(Phillips et al,2006,2008)、生态学格局(Harte et al,2008)和物种间的相互作用(Volkov et al,2009)等方面。中国也有很多用其来预测珍稀物种或入侵物种以及一些植物病虫害潜在分布区域的报道(王运生等,2007;曾辉等,2008;李明阳等,2008;马松梅等,2010),段居琦等(2011,2012)则将最大熵方法用于水稻种植区的气候适宜性研究,提出最大熵模型可以用于中国水稻种植区分布及其气候适宜性研究,但尚未分析灌溉条件下中国单季稻种植北界。
在已有研究中,确定单季稻种植北界的方法在本质上是以控制试验或对作物实际分布区域界限的调查资料为基础的,受当时试验控制条件、调查资料精度和农业生产水平影响较大,有一定局限性(程德瑜等,1994),且对气候因子综合作用下单季稻种植北 界研究较少。本研究将以黑龙江省为重点研究区域,根据单季稻种植与气候关系的最大熵模型给出的单季稻存在概率确定出中国单季稻种植的北界,并与已有结果进行对比分析,探讨该方法确定雨养和灌溉条件下中国单季稻种植北界的可行性,为黑龙江省水稻的生产布局及科学应对气候变化提供依据。 2 数据与方法 2.1 数据来源
本研究使用的中国单季稻的地理分布数据和气候数据来自于中国国家气象信息中心,包括以雨养为主的75个进行单季稻观测的农业气象观测站,1971—2000年气候标准年的中国地面气候逐日数据。其研究区域、气象台站和单季稻站点如图 1所示。
 |
| 图 1 研究区域、气象台站及进行单季稻观测的农业气象观测站Fig. 1 Location of the meteorological station,single harvest rice stations in China(a) and Heilongjiang Province(b) |
采用空间卷积算法,结合气象站点位置的截断高斯滤波算子作为距离权重方程,获取空间分辨率为10 km×10 km的日平均气温和日降水量,并将其作为基础气候数据(Thornton et al,1997; 刘宇等,2006)。经交叉验证表明,该插值方法比距离平方反比法、梯度距离平方反比法和普通克里格法等其他插值方法,所得误差小。此算法是采用最近邻域法和反向距离法的基本原理,将高斯滤波算子作为距离权重方程,并且出于计算效率的考虑,设定一个有效作用距离,即截断距离,如果一个测值点与插值目标点的距离大于算子的截断距离,则其测值对目标点的贡献为0。因此,最终的权重方程为
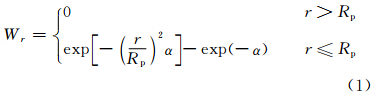
然后计算Rp作为插值目标点p的邻域截断距离,并且,用于式(1)中计算邻域内各测值点对目标点的插值权重。从而得到目标点气象要素x的插值结果
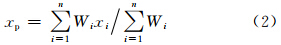
对于气温,除了平面上的空间相关外,还有随高度的变化。取气温的垂直递减率β为其平均值-0.006℃/m。经过插值站点和观测站点的高程数据修正之后的气温空间插值计算式最终为(刘宇等,2006)
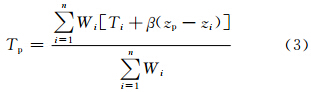
日降水量插值则在式⑵的基础上采用Thornton等(1997)的降水插值方法。 2.2 研究方法 2.2.1 气候因子计算
段居琦等(2011,2012)研究涉及到的气候因子有年降水量、湿润指数、稳定通过18℃持续日数、稳定通过10℃持续日数和≥10℃积温。
年降水量P为水分资源的绝对数量,可作为一些作物的分界线;湿润指数(IM)表示一地气候的干湿程度,反映水分收支比率,其常用的指标计算方法是降水量与蒸散量的比值。霍尔德里奇生命地带系统中的计算方法所需气候要素较少,又具有明确的生物学意义,在多个国家和地区得到应用并取得了较好的效果,所以,本文亦选用该方法


稳定通过18℃持续日数(N18):18℃是试验与实践中观测到的水稻抽穗扬花的最低温度,稳定通过18℃持续日数反映了满足水稻幼穗分化到抽穗扬花期的高温期,计算方法采用5日滑动平均法(曲曼丽,1991);稳定通过10℃持续日数(N10):10℃是喜温作物开始播种和生长的临界温度,稳定通过10℃持续日数反映了满足喜温作物生长发育的生长期,计算方法采用5日滑动平均法(曲曼丽,1991);≥10℃积温(∑T10):是喜温作物生长期内的热量累积,可用于反映喜温作物对热量的要求,采用累积计算法

最大熵模型是基于有限的已知信息对未知分布进行无偏推断的一种数学方法。该理论认为在无外力作用下,事物总是在约束条件下争取最大的自由权,在已知条件下,熵最大的事物最可能接近它的真实状态(Jaynes,1957)。最大熵统计建模就是从符合条件的分布中选择熵最大的分布作为最优分布。最大熵模型对物种存在概率的估计可以通过贝叶斯决策理论进行解释(Phillips et al,2008)。模型的目的是寻找最优的对数可能性和确定概率分布π的真实分布。
概率分布π的确定是通过研究区设定的站点X中随机选取一个站点x,如果该物种在站点x处存在,记为1,否则记为0。将响应变量(存在或不存在)设为y,那么π(x)就是受限制条件约束的概率P(x|y=1)。例如,假定物种在x点存在,则其在x点的概率为1。根据贝叶斯规则
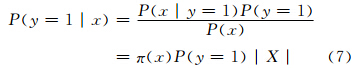
为此,对所有的x,P(x)=1/|X|。这里,P(y=1)是该物种在研究区域总的比例。P(y=1|x)是物种在x点的存在概率,对植物而言,其值是0或1,但是对散布的有机体来说,其值在0—1。式(7)显示π与存在概率相关。尽管如此,如果只有存在数据,还是不能确定该物种在研究区所占的比例。因此,不直接计算P(y=1|x),而是计算概率分布π。需要指出的是,x在这里是站点,而不是环境条件的向量。
熵最大时的概率分布满足Gibbs分布。Gibbs分布是根据特征向量权重参数化的指数分布,其形式为
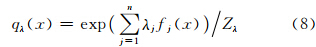
本研究在分析黑龙江省农业气候资源特征的基础上,根据单季稻种植与气候关系的最大熵模型给出单季稻在待预测区的存在概率,以确定中国单季稻种植的北界,当存在概率小于0.05时,从统计学上来看,可以认为存在的可能性很小,因而以存在概率0.05作为单季稻能否种植的界限,并考虑两种情况:水热共同作用下的单季稻种植北界和假定保证水分供应、只考虑热量条件的单季稻种植北界。以影响单季稻潜在分布的主导气候因子年降水量、湿润指数和稳定通过18℃持续日数作为环境变量,模拟水热共同作用下的单季稻种植北界,记为BJ1;不考虑水分对单季稻分布的限制,则以稳定通过18 ℃持续日数作为环境变量,模拟热量限制条件下水稻的潜在分布,记为BJ2。以单季稻种植北界为界限,量化分析单季稻种植北界的气候因子阈值。
本研究还将比较基于最大熵模型确定的单季稻种植北界与基于中国农业科学院和中国农林气候区划协作组提出的单季稻种植北界方法所确定的北界的异同,并与实际单季稻种植区对比验证,以检验该方法确定单季稻种植北界的科学性。中国农业科学院提出的单季稻种植北界是≥10℃积温≥2000℃·d,中国农林气候区划协作组提出的单季稻种植北界是稳定通过10℃持续日数≥110 d和稳定通过18℃持续日数≥30 d,分别记为BJ3和BJ4。
以上分析利用ArcGIS 9.3软件先在全中国范围内进行,然后提取黑龙江省的单季稻种植北界,进行具体分析。3 结果分析 3.1 气候因子空间分布
黑龙江省水热资源分布存在明显的区域差异(图 2)。从水分条件来看,年降水量为476—673 mm,中东部降水较多,西部和北部地区降水较少(图 2a);从热量条件来看,稳定通过18℃持续日数为4—83 d(图 2c),稳定通过10℃持续日数为80—148 d(图 2d),≥10℃积温为1178—2938℃·d(图 2e),西南部的松嫩平原和东北部的三江平原地区热量资源较丰富,其次为东南部的长白山地区,再次为北部的大、小兴安岭地区;水热共同作用下的湿润指数为0.87—1.93(图 2b),最湿润的地区是呼玛西部和漠河南部的大兴安岭地区,其次为中北部的小兴安岭地区和东南部的长白山地区,再次为三江平原地区,西南部的松嫩平原地区较干燥,湿润度最低。
 |
| 图 2 研究区域气候因子的空间分布(a.年降水量,b.湿润指数,c.稳定通过18℃持续日数,d.稳定通过10℃持续日数,e.≥10℃积温)Fig. 2 Spatial distribution of the climatic characteristics in Heilongjiang Province(a. Annual precipitation; b. Moisture index; c. Days of not less than 18 ℃ stably; d. Days of not less than 10 ℃ stably; e. Accumulated temperature of not less than 10 ℃) |
从图 3可以看出,在水热共同作用的雨养条件下,中国单季稻种植北界(BJ1)可达漠河县北部,沿漠河—塔河—呼玛中部以西的大兴安岭地区及龙江—泰来—杜尔伯特—大庆—肇州—肇源以西的地区不能种植单季稻。雨养条件下单季稻种植北界的临界气候条件是年降水量为486 mm,湿润指数为0.91—1.93,N18为8 d。相应的临界N10为95 d,临界∑T10为1448℃·d。
 |
| 图 3 中国单季稻的种植北界Fig. 3 Single harvest rice planting northern boundaries in China |
如果水稻灌溉有保障,热量限制的中国单季稻种植的北界(BJ2)则不存在,即在中国最北部漠河仍可以种植,与已有研究一致(浙江农业大学,1981);但BJ2以西的地区为水稻不能种植区。沿漠河—塔河—呼玛中部的单季稻种植界限往西略有偏移,而龙江—泰来—杜尔伯特—大庆—肇州—肇源以西的地区也能满足单季稻种植的热量需求。灌溉条件下的单季稻北界临界N10为85 d,临界∑T10为1281℃·d。
基于中国农业科学院提出方法确定的中国单季稻种植的最北端位于呼玛中部,约为52°N,北界(BJ3)为呼玛—黑河—孙吴—逊克—嘉萌—伊春—逊克—五大莲池—嫩江一线,此线以北为中国单季稻不可种植区。基于中国农林气候区划协作组提出方法确定的中国单季稻种植北界(BJ4)的最北端约位于51°N,较BJ3的最北端偏南约1个纬度;BJ4与BJ3的界限相似,但在伊春—逊克—五大连池一段,界限略偏南。本研究确定的中国单季稻种植北界(漠河北部)则分别较BJ3和BJ4的最北部偏北1.5和2.5个纬度,且其确定的单季稻可种植区也向北向西方向扩展。4 讨论与结论
Woodward(1987)指出,决定植物分布的控制因子可分为3类:(1)植物可以忍受的最低温度;(2)完成生活史所需的生长季长度和热量供应;(3)用于植物冠层形成和维持的水分供应。水稻作为喜温作物,其生长期内的最低温度不成为其种植分布的主要控制因子;水稻种植主要受改善水利设施等人为活动影响,因而水分对水稻的影响主要表现在“以水定稻”上,其主要作用是可将水稻种植区区分为雨养水稻区和灌溉水稻区。所以,最终控制水稻分布机理是完成生活史所需的生长季长度和热量供应。自然气候条件下影响单季稻分布的主导气候因子是年降水量、湿润指数和稳定通过18℃持续日数。由于在水稻生产中,对水分不足的地区经常通过灌溉来保证水分,这样影响水稻分布的主导气候因子就是稳定通过18℃持续日数。水稻要完成生活史,其关键是要保障花期免受低温冷害,确保安全齐穗(雁伍,1978)。花期低温冷害会使水稻花药发育停止,花粉发育不良,从而导致颖花空壳不育,产量下降(马树庆等,2003)。N18的主要作用是保障水稻完成从幼穗分化到抽穗扬花这一生殖生长阶段,即水稻的安全齐穗。同时,由于主导气候因子能够代表影响物种分布的气候条件,因而N18可以看作影响单季稻分布的充分必要条件。已有研究以∑T10≥2000℃·d,或者同时满足N10≥110 d和N18≥30 d作为单季稻种植界限的指标,前者可以看作是以热量供应来表示满足单季稻完成生活史的气候条件,后者是以生长季长度和热量强度持续时间来表示满足单季稻完成生命史的气候条件。
对古水稻遗存研究表明,水稻种植是由发源地向周边地区不断传播的过程,传播范围与稻作技术和气候环境关系密切(Gong et al,2007)。已有研究中确定单季稻种植北界的方法在本质上是以控制试验或对作物实际分布区域界限的调查资料为基础的,受当时试验控制条件、调查资料精度和农业生产水平影响较大,有一定局限性(程德瑜,1994)。不同的水稻品种对低温的抵抗性存在差异,势必会影响确定水稻种植界限的气候因子的阈值(王裁云,1983; 曲辉辉等,2011),如一般水稻品种在开花-灌浆期日平均气温连续3 d低于22℃时,就遭遇“寒露风”,而一些耐寒品种在花期能够忍耐日平均气温16℃的低温(张玉华,2007)。本研究根据最大熵模型给出单季稻在待预测区的存在概率,当存在概率小于0.05时,从统计学上来看,可以认为存在的可能性很小,因而以存在概率0.05作为单季稻能否种植的界限,具有明确的统计学意义。结果表明,最大熵模型方法确定的中国单季稻种植北界与已有研究结果(浙江农业大学,1981)基本一致。目前,黑龙江省的呼玛地区已经种植了单季稻(Frolking et al,2002; 朱晓禧等,2008),这表明基于中国农业科学院和中国农林气候区划协作组提出的单季稻种植北界方法由于其基于试验或调查资料,没有考虑各因子的综合作用,导致所确定的北界偏南,而基于最大熵模型方法避免了气候因子阈值选取可能存在的不确定性给确定单季稻种植界限带来较大的误差,使确定的单季稻种植北界更符合实际。
本研究表明,龙江—泰来—杜尔伯特—大庆—肇州—肇源以西的地区尽管水分不足,但热量充足(图 2),能够满足单季稻生产。该地区已经成为松嫩平原的水稻生产区。本研究给出的BJ1体现了雨养(水热共同作用)下单季稻种植的北界,其以东以南地区是以雨养为主的单季稻种植区;BJ2体现了灌溉条件(热量制约)下的单季稻种植北界,其以东以南地区是灌溉和雨养条件下的单季稻可种植区;BJ1和BJ2之间的地区,是以灌溉措施保障单季稻水分需求为主的单季稻可种植区。
气候系统是一个复杂系统,存在年际、年代际变率,极端天气、气候还常造成气象灾害(王绍武等1999;张庆云等,2008)。气候变化也会影响作物种植结构(Wang,1997)。本研究给出的是一个气候标准年(1971—2000年)的中国单季稻种植北界,未考虑单季稻种植可能存在的农业气象灾害风险;在当前全球变化背景下,中国单季稻种植北界在年际、年代际是否有变化及怎样变化,尚需要进一步研究。
| 程德瑜, 高安, 古建泉. 1994. 农业气候学. 北京: 气象出版社, 13-26 |
| 程叶青, 张平宇. 2005. 中国粮食生产的区域格局变化及东北商品粮基地的响应. 地理科学, 25(5): 513-520 |
| 段居琦, 周广胜. 2011. 中国水稻潜在分布及其气候特征. 生态学报, 31(22): 6659-6668 |
| 段居琦,周广胜. 2012. 我国单季稻种植区的季候适宜性.应用生态学报,23(2):426-432 |
| 江爱良. 1982. 论我国水稻的种植上限. 地理科学, 2(4): 291-301 |
| 李明阳, 巨云为, Kumar S, et al. 2008. 美国大陆外来入侵物种斑马纹贻贝 (Dreissena polymorpha) 潜在生境预测模型. 生态学报, 28(9): 4253-4258 |
| 刘宇, 陈泮勤, 张稳等. 2006. 一种地面气温的空间插值方法及其误差分析. 大气科学, 30(1): 146-152 |
| 马树庆, 王琪, 沈享文等. 2003. 水稻障碍型冷害损失评估及预测动态模型研究. 气象学报, 61(4): 507-512 |
| 马松梅, 张明理, 张宏祥等. 2010. 利用最大熵模型和规则集遗传算法模型预测孑遗植物裸果木的潜在地理分布及格局. 植物生态学报, 34(11): 1327-1335 |
| 曲辉辉, 姜丽霞, 朱海霞等. 2011. 孕穗期低温对黑龙江省主栽水稻品种空壳率的影响. 生态学杂志, 30(3): 489-493 |
| 曲曼丽. 1991. 农业气候分析方法30例. 北京: 北京农业大学出版社, 1-9 |
| 王裁云. 1983. 滇中水稻冷害的强低温长期预报. 气象学报, 41(3): 305-312 |
| 王绍武, 朱锦红. 1999. 国外关于年代际气候变率的研究. 气象学报, 57(3): 376-384 |
| 王运生, 谢丙炎, 万方浩等. 2007. 相似穿孔线虫在中国的适生区预测. 中国农业科学, 40(11): 2502-2506 |
| 邢丁亮, 郝占庆. 2011. 最大熵原理及其在生态学研究中的应用. 生物多样性, 19(3): 295-302 |
| 雁伍. 1978. 水稻的安全齐穗期. 气象, (7): 29-31 |
| 云雅如, 方修琦, 王丽岩等. 2007. 我国作物种植界线对气候变暖的适应性响应. 作物杂志, (3): 20-23 |
| 曾辉, 黄冠胜, 林伟等. 2008. 利用MaxEnt预测橡胶南美叶疫病菌在全球的潜在地理分布. 植物保护, 34(3): 88-92 |
| 张庆云, 陶诗言, 彭京备. 2008. 我国灾害性天气气候事件成因机理的研究进展. 大气科学, 32(4): 815-825 |
| 张新时, 杨奠安, 倪文革. 1993. 植被的PE(可能蒸散) 指标与植被气候分类(三):几种主要方法与PEP程序介绍. 植物生态学与地植物学学报, 17(2): 97-109 |
| 张学文. 1987a. 云滴的表面自由能统计分布理论. 新疆气象, (4): 7-11 |
| 张学文. 1987b. 最大熵原理及其在气象中的应用. 新疆气象, (7): 2-8, 38 |
| 张学文, 马力. 1995. 熵气象学简介. 气象, 21(1): 52-56 |
| 张谊光. 1983. 我国不同气候生态型水稻品种的种植上限及其温度条件. 资源科学, (2): 65-72 |
| 张玉华. 2007. 北方寒地耐冷抗病优质高产新品种龙稻5号. 中国种业, (8): 73 |
| 浙江农业大学. 1981. 实用水稻栽培学. 上海: 上海科学技术出版社, 2-5 |
| 郑国光. 1988. 对熵原理得出的理论云滴谱方程的验证. 新疆气象, (2): 45-47, 56 |
| 中国农林作物气候区划协作组. 1987. 中国农林作物气候区划. 北京: 气象出版社, 36-55 |
| 中国农业科学院. 1986. 中国稻作学. 北京: 农业出版社, 85-122 |
| 周家斌, 黄嘉佑. 1997. 近年来中国统计气象学的新进展. 气象学报, 55(3): 297-305 |
| 朱晓禧, 方修琦, 王媛. 2008. 基于遥感的黑龙江省西部水稻、玉米种植范围对温度变化的响应. 地理科学, 28(1): 66-71 |
| Frolking S, Qiu J J, Boles S, et al. 2002. Combining remote sensing and ground census data to develop new maps of the distribution of rice agriculture in China. Global Biogeochem Cycle, 16(4): 1091, doi: 10.1029/2001GB001425 |
| Gong Z T, Chen H Z, Yuan D G, et al. 2007. The temporal and spatial distribution of ancient rice in China and its implications. Chin Sci Bull, 52(8): 1071-1079 |
| Harte J, Zillio T, Conlisk E, et al. 2008. Maximum entropy and the state-variable approach to macroecology. Ecology, 89(10): 2700-2711 |
| Jaynes E T. 1957. Information theory and statistical mechanics. Physical Review, 106(2): 620-630 |
| Phillips S J, Anderson R P, Schapire R E. 2006. Maximum entropy modeling of species geographic distributions. Ecol Mod, 190(3-4): 231-259 |
| Phillips S J, Dudík M. 2008. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography, 31(2): 161-175 |
| Shipley B. 2010. Community assembly, natural selection and maximum entropy models. Oikos, 119(4): 604-609 |
| Thornton P E, Running S W, White M A. 1997. Generating surfaces of daily meteorological variables over large regions of complex terrain. J Hydrol, 190(3-4): 214-251 |
| Volkov I, Banavar J R, Hubbell S P, et al. 2009. Inferring species interactions in tropical forests. Proc Natl Acad Sci USA, 106(33): 13854-13859 |
| Wang F T. 1997. Impacts of climate change on cropping system and its implication for agriculture in China. Acta Meteor Sinica, 11(4): 407-415 |
| Woodward F I. 1987. Climate and Plant Distribution. Cambridge: Cambridge University Press, 62-107 |