
文章信息
- 罗天同, Gabriel YEDID
- LUO Tiantong, Gabriel YEDID
- 基于数字进化研究大灭绝对群落稳定性和协调停滞现象出现的影响
- Effects of mass extinction on community stability and emergence of coordinated stasis with digital evolution
- 南京农业大学学报, 2018, 41(1): 88-97
- Journal of Nanjing Agricultural University, 2018, 41(1): 88-97.
- http://dx.doi.org/10.7685/jnau.201703016
-
文章历史
- 收稿日期: 2017-03-16




“协调停滞”的概念最早是Brett等[1]对志留纪—泥盆纪时期的北阿拉巴契亚盆地中动物区系(fauna)的观察研究而提出的:在11个被大灭绝事件分隔的生态-进化子单元(ecological-evolutionary subunit)中, 大多数物种停留在其偏爱的环境中, 使得生物相(biofacies)长时间保持稳定。协调停滞现象在一定程度上是间断平衡理论(punctuated equilibria)的延伸:大多数物种在较长地质历史时期中发生的进化改变是极其微小的[2]。与间断平衡理论不同的是, 协调停滞现象强调的是大量谱系同时出现停滞和改变的现象[1]。协调停滞现象在生态系统中具体体现在以下3个方面:1)谱系的停滞; 2)生物相保持稳定; 3)谱系与生物相同时发生改变(turnover)[3]。有关协调停滞现象出现的机制, 研究者提出了数个仍存在争议的假说, 其中2个最重要的是栖息地追踪(habitat tracking)[4]和生态封锁(ecological locking)[5]。栖息地追踪的理论认为物种总会追踪它们偏好的栖息地, 而生态封锁描述的是群落内复杂的种间关系阻碍了整个群落的进化。
自然界中生态群落的稳定性通常由群落结构的多样性(物种丰富度)、物种构成(species composition)以及物种间相互作用的模式决定[6]。通常可以通过以下几个方面对群落稳定性进行验证和评估:1)恒定性(constancy)和持久性(persistence):群落各方面的生态属性长时间保持不变的能力; 2)抵抗力(resistance):群落以及生态系统承受干扰的能力; 3)恢复力(resilience):群落在干扰之后恢复到参照状态的速度; 4)稳定状态(stable state)和多稳态(alternative stable state):群落在经历干扰后是否展现出恢复到原始状态或者转变到另一个替代稳定状态的趋势。本研究将对群落不同方面的稳定性以及优势生态型如何保持地位进行研究说明。
数字进化是一种利用软件研究进化理论的方法, 其建立的虚拟模型与真实生物群落有以下共同特征:通过复制进行繁殖; 由突变产生变化; 个体之间的繁殖率不同; 由可耗减的资源产生对个体的选择压力。与传统的试验进化生物学以及古生物学的研究方法相比, 数字进化具有更高的重复性、透明度、计算精度以及更强的统计学功能。本研究中所使用的数字进化工具Avida是一个被广泛应用的开源软件平台(http://avida.devosoft.org)。该平台已被成功地应用于多个进化生物学领域的研究中。例如:复杂性状的进化[7]、高突变率下的进化过程[8]、生态系统的形成及多样化[9]、性选择[10]以及有害突变在进化中的作用[11]等, 相关研究成果[8, 12-14]发表在包括《Science》和《Nature》在内的数十个高水平的学术杂志上。
在本研究中, 我们运用Avida平台的优势, 结合多种数据分析工具和方法, 对不同强度和类型大灭绝条件下的数字群落在不同水平上的稳定性和协调停滞现象的出现条件及普遍性进行验证和评估, 为相关古生物学及试验进化生物学的相关研究提供了参考和依据。
1 材料与方法 1.1 试验平台本研究工作使用的主要研究工具为Avida数字进化平台。所有的试验实施以及数据分析均在一个包含6个计算节点、36核的计算机集群上完成(每个计算节点的硬件配置为:Intel i7-4930(3.4 GHz)CPU, 16 GB内存, 500 GB硬盘)。主要数据分析及作图工具为MATLAB(R2010a)和R(3.2.3)。
1.2 环境配置环境配置包括2个主要特征:多种可耗减资源以及互养(cross-feeding)的生态互作等级制度(图 1)。资源的可耗减性即数字个体(Avidians)对某种资源的利用会降低该资源对其他个体的可利用性, 包括利用该资源获得的收益, 这种资源竞争模型类似于在微生物中出现的竞争模式。具有相同资源需求的个体之间的竞争导致它们提高对资源的利用率, 并且对资源的要求重叠最少的个体很可能共存[9, 14]。在互养的环境中, 只有有限种类的资源是外部供给的, 其余的资源将以副产物的形式产生:当Avidians成功地执行有关指令以利用某些资源后, 将产生一定数量和种类的副产物, 这些副产物将成为其他Avidians的可利用资源。这种设置将通过促进生态位构建(niche construction)来引发基本营养结构的进化[15]。由于缺乏捕食关系的设定, 本研究中的互养关系不能被称为真正的食物网, 但这种营养结构已经在之前的微生物进化试验中被发现[16-17], 因此该模型可用于研究生物进化的一般规律。

|
图 1 互养关系示意图[18] Figure 1 Cross-feeding relationships 箭头的颜色表示不同的营养级(绿色:初级产物; 粉色:一级消费者; 品红色:次级消费者; 红色:顶级消费者)。R1~R9为9种资源。NOT、NAND、AND、ORN、OR、ANDN、NOR、XOR和EQU为对应的9种利用资源的功能。下同。 Arrows are color coded by trophic levels(green:production; pink:primary consumer; magenta:secondary consumer; red:top consumer).R1-R9 are 9 resources. NOT, NAND, AND, ORN, OR, ANDN, NOR, XOR and EQU are 9 corresponding functions that consume the resources. The same as follows. |
本研究中所有试验重复都是由一个仅具有自我复制能力的手动编译祖先开始的, 它的后代将会进化出执行特定计算功能的能力, 当其形成一个可以有效利用资源的群落后[19-21], 将被用于之后的一系列试验。Avida中的绝对时间是用“update”来衡量的, Avidians在一个单位“update”里执行大约30条指令, 通常一个世代需要经历5至10个update, 具体时长将由Avidians表现出的复杂性决定。
本试验设计将引入2个重要的进化历程:大灭绝以及灭绝后的恢复过程。具体的大灭绝以及恢复过程将通过5个试验条件(试验对照、强压力灭绝、弱压力灭绝、强脉冲灭绝和弱脉冲灭绝)来实现, 与之前的研究[15]类似, 每种试验条件进行100次重复(每种条件下的试验重复编号均为1 000, 1 100, 1 200, …, 10 900, 共100个)(图 2)。

|
图 2 大灭绝及恢复过程的设置 Figure 2 The setups of mass extinction and recovery |
每个试验重复在没有大灭绝加入的情况下运行205 000个update。
1.3.2 强脉冲灭绝(strong pulse)每个试验重复首先运行100 000个update, 之后对群落实施一次大量的瞬时个体移除:幸存者被随机地从群落中选出, 存活率为整个群落中个体数量的0.1%。随后群落经历100 000个update的恢复过程, 条件与灭绝前相同。
1.3.3 弱脉冲灭绝(weak pulse)灭绝前、后的各100 000个update均与以上条件相同, 但是存活率为整个群落中个体数量的1%。
1.3.4 强压力灭绝(strong press)灭绝前的100 000个update与以上条件相同, 之后将进入强压力灭绝时期:在之后5 000个update的时间里, 资源的流入量将降低2个数量级, 随后恢复正常。
1.3.5 弱压力灭绝(weak press)灭绝前、后的各100 000个update以及弱压力灭绝时期均与强压力灭绝相同, 但是弱压力灭绝时期的资源流入量是强压力灭绝的10倍。
1.3.6 时间穿越入侵试验(time-shift invasion trials)该试验是用来验证停滞的出现以及群落在表现水平(performance level)稳定性的主要方法, 其可以验证稳定的生态型达到优势地位和组建群落的“生态核心(ecological core)”是否是通过“红皇后(Red Queen)”过程[22]达到的, 即任何个体发生的有利进化改变会降低群落中其他个体的适合度。简单来说, 时间穿越入侵试验即为同一试验重复中不同时期群落成员的竞争试验(图 3)。具体的实施方法有以下3种:1)完整群落竞争试验(full community competitions):将所有试验条件实施前(pre-treatment, PT)的物种及所有试验结束时(end-experiment, EE)的物种放在一起竞争。2)替代试验(replacement trials):分别用每个EE物种替代掉与之具有相同表型的PT物种。3)入侵试验(invasion trials):如果2个群落的物种数量不等, 或者无法将所有物种互相配对, 则分别将每个EE物种直接置入PT群落。类似地, 也可以将PT物种分别置入EE群落中, 我们将此称为“逆向入侵试验(inverse invasion trials)”。

|
图 3 时间穿越入侵试验示意图 Figure 3 The diagram of time-shift invasion trials |
对时间穿越入侵试验结果的预测如下:1)真正的停滞:EE群落的物种和PT群落的物种相比并没有明显的进步, 双方均在约50%的试验重复中展现出优势; 2)完全过渡性进化(fully transitive evolution):EE群落的物种总是优于PT群落的物种; 3)完全非过渡性进化(fully intransitive evolution):PT群落的物种总是优于EE群落的物种; 4)部分非过渡性进化(partially intransitive evolution):入侵试验形成的群落既包含了PT群落物种, 也包含了EE群落物种。
1.4 数据分析 1.4.1 平均级转移(mean rank shift, MRS)MRS是一个用来从不同的时间节点评估群落变化的物种特征敏感(identity-sensitive)指数[23], 其可以反映出群落的生态组成结构随时间的变化。我们使用群落中10个多度最高的物种, 将统计值定义为MRS10。其计算方法如下[17]:
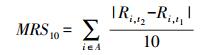
|
式中:t1和t2是2个被调查的时间点(在本研究中即PT和EE); A是在t1和t2时间点上多度最高的10个物种的合集; Ri, tx是物种i在时间点tx时的相对级(relative rank)。当研究对象为PT时的优势种时, MRS10描述的是这些物种在大灭绝后的命运, 这种研究思路称为“前瞻视角(prospective view)”; 而当研究对象是t2时的优势种时, 其描述的是这些物种在t1时的等级, 也就是EE群落的形成过程, 这种思路称为“回顾视角(retrospective view)”。MRS10可以被视为一个群落的相似性衰减指数(decay of community similarity index), 或者是群落的差异性增长指数(increase of community dissimilarity index), 即MRS10值越高, 则该时间点的群落与参考群落之间的相似性越低, 或者说差异性越高。
1.4.2 互熵(mutual entropy)与分工(division of labor)分工是社会性(sociality)的最重要属性之一。在本研究中, 我们借鉴Gorelick等24利用Shannon[25]的信息论(information theory)中的互熵、边缘熵(marginal entropy)以及香农多样性指数(Shannon′s DIT)的特化度(specialization)作为衡量分工的指标, 分工的量化有助于我们了解群落在不同时间点状态的改变。其计算方法如下:

|
式中:X和Y分别是个体与任务的合集; p(x, y)为X和Y的联合概率分布函数(joint probability distribution function); p(x)和p(y)分别是X和Y的边缘概率分布函数(marginal probability distribution function); DY|X即DIT; I为互信息(mutual information); H为熵(entropy)。9种不同的功能就相当于分工中的任务, 表达功能的Avidians就相当于分工中的个体。若DY|X的值趋于0, 说明群落中的个体分工更加“专门化(specialized)”, 即每个个体表达的功能数较少且物种数量相对较多; 若DY|X的值趋于1, 说明群落中的个体分工更加“均质化(generalized)”, 即每个个体表达的功能数较多且物种数量相对较少。
1.4.3 MRS10和DIT的时间序列数据的断点分析(breakpoint analyses)我们借鉴了针对真实生态系统中时间序列数据的断点分析方法[26], 使用R语言中的strucchange程序包(https://www.r-project.org/)检验MRS10和DIT的时间序列数据的结构变化并将结果进行可视化处理, 以验证它们之间改变的协调性及同步性。
1.4.4 时间穿越入侵试验结果的分析方法我们使用与微生物试验更加相似的瞬时出生率(instantaneous birth rate, IBR)作为适合度的衡量指标。做出IBR的时间序列曲线之后, 分别利用赤池信息准则(Akaike information criterion, AIC)以及似然比检验(Reitan′s likelihood ratio test)确定曲线最初增长阶段(initial increase)以及平稳状态(equilibrium fitness)在有、无入侵者情况下的差异显著性, 从而检验入侵者对群落其他个体的影响, 以确定群落稳定性的变化以及是否有协调停滞现象的出现。
2 结果与分析 2.1 平均级转移我们以MRS10不大于0.1作为生态稳定群落的判断标准, 将所有试验重复分为3类:极度不稳定的群落、一般群落和非常稳定的群落(表 1)。从表 1可知:对群落的生态稳定性影响最大的试验条件为强压力灭绝。在强压力灭绝的条件下, 几乎没有任何群落表现出较高的稳定性。也就是说经历了强压力灭绝后, 群落的物种等级发生了极大的逆转, 大部分PT优势种被大灭绝淘汰, 取而代之的是等级较低的物种甚至是大灭绝后出现的新种。这种现象的出现是由于在资源充足的条件下的优势种并不能够很好地适应资源匮乏的环境, 低等级的物种趁机占据了空缺的生态位, 从而建立优势。
| 试验重复 Replicates |
非常稳定群落数 Number of very stable communities |
一般群落数 Number of average cases |
极度不稳定群落数 Number of highly unstable communities |
|
| 试验对照 Control |
PROSP RETRO |
30 29 | 60 64 | 10 7 |
| 弱压力灭绝 Weak press |
PROSP RETRO |
19 24 | 74 63 | 7 13 |
| 弱脉冲灭绝 Weak pulse |
PROSP RETRO |
32 34 | 63 63 | 5 3 |
| 强压力灭绝 Strong press |
PROSP RETRO |
1 0 | 52 41 | 47 59 |
| 强脉冲灭绝 Strong pulse |
PROSP RETRO |
7 15 | 82 67 | 11 18 |
| 注: PROSP:前瞻视角Prospective view; RETRO:回顾视角Retrospective view.以上平均级转移结果均以每100个群落计。The MRS10 results are calculated by 100 communities. | ||||
对群落稳定性影响较大的试验条件是强脉冲灭绝。强脉冲灭绝造成的极度不稳定群落数量比强压力灭绝少了很多, 大部分的群落表现出了一定的稳定性。然而, 从两个不同的视角进行研究可知, 在强脉冲条件下回顾视角方法认定的稳定群落数量比前瞻视角多了1倍以上。这种现象是由于强脉冲的随机性导致的。随机移除个体可能会导致等级最高的优势物种灭绝, 而等级较低的优势物种存活下来, 取而代之成为最优物种。从回顾视角来看, 试验结束时等级最高的优势物种并没有经历特别大的等级逆转, 因此群落表现出了较高的稳定性。
弱压力灭绝也对群落的稳定性造成了一定程度的影响, 但相比强灭绝条件来说, 其造成的影响弱了很多。弱脉冲灭绝几乎没有对群落的稳定性产生任何影响, 甚至其造成的极不稳定的群落数比试验对照组还少。这同样也是由脉冲灭绝的随机性造成的, 某些群落中将要发生等级逆转的低等级物种恰好在脉冲灭绝的条件下被移除了, 从而使高等级物种的优势得以保持。
综合4种灭绝条件以及试验对照, 在总共100个群落中有3个群落表现出了较高的生态稳定性, 分别是群落X3000、X4200和X5000。它们在除了强压力灭绝之外的所有试验条件下均保持了相当高的生态稳定性(图 4)。

|
图 4 3个稳定群落的MRS10结果 Figure 4 The MRS10 results of the 3 stable communities 1)X3000:群落3000 Community 3000;X4200:群落4200 Community 4200;X5000:群落5000 Community 5000。2)左侧为前瞻视角, 右侧为回顾视角。Left figures:Prospective views; Right figures:Retrospective views. 3)图中水平虚线为0.1。The horizontal dash lines are 0.1. |
比较不同试验条件下大灭绝后群落与大灭绝之前群落的DIT, 以及不同试验条件下大灭绝后群落与试验对照的DIT, 并且做出所有100个试验重复差异的正态分布直方图(图 5)。无论是以灭绝前群落还是以灭绝后的试验对照群落作为参照, 越强的灭绝条件会对群落的DIT值产生越大的影响, 使DIT差异的波动范围变得越大, 并且压力灭绝相对于脉冲灭绝来说对群落的影响更大, 与MRS10的结果相吻合。

|
图 5 所有试验条件DIT差异的正态分布立体直方图 Figure 5 The 3D histograms of the distributions of the DIT differences for all treatments |
通过每个单独试验重复中群落在不同的灭绝条件下的DIT与试验对照DIT差异的时间序列曲线, 并对其进行断点分析和差异显著性分析, 找出DIT稳定性较高的群落。按照稳定群落DIT不大于0.1的判断标准, 在100组试验重复中, 稳定性较高、中等及较低的群落数量分别为11、40和49。最后通过生成不同灭绝条件下所有试验重复的DIT时间序列曲线来消除由于DIT变化趋势造成的误差, 最终得到3个DIT稳定群落:群落3000、4200和9600(图 6), 它们在除了强压力灭绝之外的其他条件下均保持了较高的DIT稳定性。

|
图 6 稳定群落灭绝后DIT与试验对照DIT差异的时间序列曲线 Figure 6 The time series data of the DIT differences between EE stable communities and control |
没有出现完全的停滞以及过渡性进化, 并且几乎所有的试验对照(98/100)、弱灭绝条件(弱压力灭绝96/100, 弱脉冲灭绝98/100)的试验重复都属于完全的过渡性进化, 只有强灭绝条件的试验重复出现了一定数量的非过渡性进化的例子(表 2)。即使是在展现出一定非过渡性的试验重复中, 大部分PT群落的物种在竞争结束时的适合度还是有明显下降, 说明对于单个物种来说, 即使是展现出完全的非过渡性, 其余的EE群落物种还是能对其造成相当大的影响。此外, 还有一部分非过渡性进化是由群落的生态完整性(eco-complete community)造成的, 即某1或2个群落中有空缺的生态位, 使得另一群落中占有该生态位的物种更容易在竞争中获胜。
| 指标 Indes |
试验对照 Control |
弱压力灭绝 Weak press |
弱脉冲灭绝 Weak pulse |
强压力灭绝 Strong press |
强脉冲灭绝 Strong pulse |
| 最终群落的平均物种数量 Average number of ecotypes in final community |
5.97 | 6.08 | 5.99 | 6.45 | 6.12 |
| 最终群落中来自PT群落的的平均物种数量 Average number of ecotypes from PT community |
0.02 | 0.06 | 0.04 | 1.62 | 0.63 |
| 最终群落中来自EE群落的平均物种数量 Average number of ecotypes from EE community |
5.95 | 6.02 | 5.95 | 4.83 | 5.49 |
| PT及EE群落均为生态完整的试验重复数量 Number of replicates where both communities eco-complete |
85 | 85 | 84 | 56 | 74 |
| 展现非过渡性的试验重复数量 Total number of replicates showing intransitivity |
2 | 4 | 2 | 56 | 40 |
| 展现非过渡性的生态完整试验重复数量 Total number of eco-complete replicates showing intransitivity |
2 | 4 | 2 | 40 | 25 |
| 竞争降低PT群落物种适合度的试验重复数量 Number of carryovers with lower fitness vs. native |
2 | 4 | 2 | 34 | 37 |
| 竞争增加PT群落物种适合度的试验重复数量 Number of carryovers with higher fitness vs. native |
0 | 0 | 0 | 11 | 2 |
| 竞争几乎没有影响PT群落物种适合度的试验重复数量 Number of carryovers with about same fitness as native |
0 | 0 | 0 | 11 | 1 |
5种试验条件下共500组试验重复中, 有65组满足进行替代试验的要求(试验对照20组, 强压力灭绝1组, 强脉冲灭绝4组, 弱压力灭绝19组, 弱脉冲灭绝21组)。在这65组替代试验中, 只有3组群落展现出了部分物种的长期停滞现象, 即EE代替者没有对其他PT物种产生显著的影响(试验对照:2000、3100;强压力灭绝:3600)。其中, 试验对照的2个群落中分别只有1组物种处于停滞状态, 而强压力灭绝的一个群落中则有3组物种处于停滞。大部分群落的替代试验的结果都显示群落处于长期的完全过渡性进化中, 特别被证明具有较高生态稳定性的群落, 它们的群落成员在“表现”层面上并不具有同样的稳定性(图 7)。来自EE群落的替代者都对PT群落成员产生了影响, 这种影响的范围很广(0.87%~1161%), 并且几乎都是负面的, 使PT群落成员的适合度下降。图 7为来自分工极端专门化的群落3000(9个物种分别表达9种功能), 从图中可见, 不同入侵者对PT群落物种的选择系数随时间不断增长, 并没有保持停滞。

|
图 7 群落3000的选择系数随时间的变化 Figure 7 The change of selection coefficient in community 3000 |
除了可以用来进行替代试验的群落外, 大部分的群落都被用来进行入侵试验, 它们中出现了较多个体水平停滞的例子(表 3)。在入侵试验中出现“表现”水平停滞试验重复的比例远远多于替代试验, 并且灭绝强度越大, 出现停滞的概率越大, 停滞表现的越明显, 但并没有完整群落的协调停滞现象出现。这样的结果恰恰与群落生态稳定性的试验结果相反。也就是说, 生态稳定性越高, 物种构成越稳定的群落, 不同时期的物种在“表现”水平上出现停滞的概率就越低。这种现象说明了大灭绝后出现的新物种不一定比灭绝前与其生态地位及作用相当的物种在竞争中更具优势。许多EE群落物种作为入侵者并不能够成功入侵到PT群落当中, 但却可以在完整的群落竞争试验中存活下来, 说明物种的适合度是与群落结构高度相关的(community context-dependent), 在某一时间点群落中高适合度的物种并不一定能够在其他时间点的群落中表现出同样水平的优势。
| 指标Index | 试验对照 Control |
弱脉冲灭绝 Weak pulse |
弱压力灭绝 Weak press |
强脉冲灭绝 Strong pulse |
强压力灭绝 Strong press |
| 出现“表现”水平停滞的试验重复数占总试验重复数的比例 The proportion of replicates showing stasis |
4/80 | 7/79 | 4/81 | 26/96 | 41/99 |
| 每个独立入侵试验中对PT群落无显著影响的平均入侵者数 Average invaders that have no significant effects on residents |
1.25 | 1.14 | 1 | 1.42 | 2 |
| 停滞现象最显著的群落入侵试验中对PT群落无显著影响的入侵者数占总入侵者数的比例 The proportion of invaders that have no significant effects on residents in most stable community |
2/7 | 2/8 | 1/5 | 4/6 | 5/7 |
参照完整的群落竞争试验的结果, 对群落出现非过渡性的试验重复进行逆向入侵试验。其结果证明了PT物种在完整群落竞争试验中展现出优势的真实性, 即其在逆向入侵试验中相对与其相匹配的EE物种同样具有优势。
3 讨论古生物学家很早就发现, 在地球生命的演化历史中, 会常规性地发生大灭绝事件, 这些大灭绝发生的频率不高, 但往往是毁灭性的, 并导致物种多样性、形态多样性以及生态结构在极短的地质学时间内发生巨大变化[27]。同时, 大灭绝也是一个重要的进化动力:淘汰长期的优势种并且给等级较低的物种提供进化和多样化的空间。
在本研究中, 我们利用了Avida中的数字化个体作为试验对象来探讨大灭绝对群落各个水平的稳定性和停滞现象的影响, 通过设置与古生物学研究结果相关的灭绝条件[28]以及利用其在数据处理和重复试验能力中的优势为古生物学和传统试验进化生物学方法提供了补充。结果显示:多数试验对照群落保持了较低的MRS10和较稳定的DIT, 说明群落中由优势物种组成的“生态核心”长时间保持稳定。数字群落这样的特点与真实生物群落具有的丰度高和持久性强的特点[29]保持一致。大灭绝对多数群落的稳定性影响都不是长期和实质性的, 群落经过短时间内的剧烈改变后倾向于恢复到稳定状态, 但群落的物种构成和多样性程度则可能在很长一段时间内无法恢复; 并且, 灭绝的强度越大, 群落受到的影响和恢复的难度越大。上述结果与之前的数字进化[30]及古生物学研究结果[23, 31]一致。相比经历大灭绝的群落, 试验对照组的群落在生态水平表现出了较高的稳定性, 然而在“表现”水平上, 几乎所有的试验对照组群落的进化过程都属于完全的过渡性, 反而大灭绝使群落表现出了部分的非过渡性。上述结果说明大灭绝对群落在各个水平稳定性的影响均不相同, 并且群落表现出的停滞现象均为非协调的(uncoordinated)。
本研究使用与真实微生物系统更加相似的物种瞬时出生率作为适合度的衡量单位。试验结果显示物种适合度的变化符合“红皇后”过程:环境属性很大程度上由个体间的相互作用决定, 而某一物种或者个体对相互作用的改变必然会导致其他物种或个体的改变[21], 即任何物种或个体发生有利的进化改变必然会降低其他物种或个体的适合度。此结果揭示了以互养关系为基础的试验系统是导致群落协同进化的根本原因, 缺乏更加复杂的种间相互作用, 使得物种的进化改变很大程度上是被其他物种已经发生的改变所影响。
本研究中物种及个体之间的相互作用过于简化, 在未来的研究中, 我们将移入包含梯度资源的复杂系统, 并且允许数字化个体对可移动资源进行追踪, 从而更符合真实个体间的相互作用, 例如捕食[32]和寄生等, 并且可以将种间相互作用的稳定性作为衡量群落稳定性的另一个标准[33]。这些新特性的引入将会弥补此前研究中环境异质性参数设定的不足。
| [1] | Brett C E, Baird G C. Coordinated stasis and evolutionary ecology of Silurian to Middle Devonian faunas in the Appalachian Basin[M]. New York: Columbia University Press, 1995, 285-315. |
| [2] | Eldredge N, Gould S J. Punctuated equilibria:an alternative to phyletic gradualism[M]. San Francisco: Freeman, 1972, 82-115. |
| [3] | Brett C E. Coordinated stasis reconsidered:a perspective at fifteen years[M]. New York: Springer, 2012, 23-36. |
| [4] | Brett C E, Mclaughlin P I. Response of shallow marine biotas to sea-level fluctuations:a review of faunal replacement and the process of habitat tracking[J]. Palaios, 2014, 22(22): 228-244. |
| [5] | Morris P J, Ivany L C, Schopf K M, et al. The challenge of paleoecological stasis:reassessing sources of evolutionary stability[J]. Proc Natl Acad Sci USA, 1995, 92(24): 11269-11273. DOI: 10.1073/pnas.92.24.11269 |
| [6] | Ives A R, Carpenter S R. Estimating community stability and ecological interactions from time-series data[J]. Ecological Monographs, 2003, 73(2): 301-330. DOI: 10.1890/0012-9615(2003)073[0301:ECSAEI]2.0.CO;2 |
| [7] | Goldberg D E. The design of innovation:lessons from and for competent genetic algorithms[M]. Berlin: Springer Science and Business Media, 2013, 107-110. |
| [8] | Wilke C O, Wang J L, Ofria C, et al. Evolution of digital organisms at high mutation rates leads to survival of the flattest[J]. Nature, 2001, 412(6844): 331-333. DOI: 10.1038/35085569 |
| [9] | Cooper T F, Ofria C. Evolution of stable ecosystems in populations of digital organisms[M]. Cambridge: MIT Press, 2002, 227-232. |
| [10] | Chandler C H, Ofria C, Dqorkin I. Runaway sexual selection leads to good genes[J]. Evolution, 2013, 67(1): 110-119. DOI: 10.1111/evo.2013.67.issue-1 |
| [11] | Covert A W, Lenski R E, Wilke C O, et al. Experiments on the role of deleterious mutations as stepping stones in adaptive evolution[J]. Proc Natl Acad Sci USA, 2013, 110(34): E3171-E3178. DOI: 10.1073/pnas.1313424110 |
| [12] | Lenski R E, Ofria C, Collier T C, et al. Genome complexity, robustness and genetic interactions in digital organisms[J]. Nature, 1999, 400(6745): 661-664. DOI: 10.1038/23245 |
| [13] | Lenski R E, Ofria C, Pennock R T, et al. The evolutionary origin of complex features[J]. Nature, 2003, 423(6936): 139-144. DOI: 10.1038/nature01568 |
| [14] | Chow S S, Wilke C O, Ofria C, et al. Adaptive radiation from resource competition in digital organisms[J]. Science, 2004, 305(5680): 84-86. DOI: 10.1126/science.1096307 |
| [15] | Odling-Smee F J, Laland K N, Feldman M W. Niche construction:the neglected process in evolution[M]. Princeton: Princeton University Press, 2003. |
| [16] | Rozen D E, Schneider D, Lenski R E. Long-term experimental evolution in Escherichia coli:Ⅷ. Phylogenetic history of a balanced polymorphism[J]. Journal of Molecular Evolution, 2005, 61(2): 171-180. DOI: 10.1007/s00239-004-0322-2 |
| [17] | Robling M, Mcnamara R, Bennert K, et al. The effect of the talking diabetes consulting skills intervention on glycaemic control and quality of life in children with type 1 diabetes:cluster randomised controlled trial(DEPICTED study)[J]. BMJ, 2012, 344: e2359. DOI: 10.1136/bmj.e2359 |
| [18] | Yedid G, Heier L. Effects of random and selective mass extinction on community composition in communities of digital organisms[M]. Berlin Heidelberg: Springer, 2012, 43-64. |
| [19] | Grabowski L M, Bryson D M, Dyer F C, et al. Early evolution of memory usage in digital organisms[C]//Proceedings of the 12th International Conference on the Synthesis and Simulation of Living Systems. Cambridge: MIT Press, 2010: 224-231. https://www.researchgate.net/publication/228868397_Early_Evolution_of_Memory_Usage_in_Digital_Organisms |
| [20] | Grabowski L M, Bryson D M, Dyer F C, et al. Clever creatures: case studies of evolved digital organisms[C]//Proceedings of the Eleventh European Conference on the Synthesis and Simulation of Living Systems, 2011: 276-283. https://www.researchgate.net/publication/232808309_Clever_Creatures_Case_Studies_of_Evolved_Digital_Organisms |
| [21] | Grabowski L M, Bryson D M, Dyer F C, et al. A case study of the de novo evolution of a complex odometric behavior in digital organisms[J]. PLoS ONE, 2013, 8(4): e60466. DOI: 10.1371/journal.pone.0060466 |
| [22] | Liow L H, van Valen L, Stenseth N C. Red queen:from populations to taxa and communities[J]. Trends in Ecology and Evolution, 2011, 26(7): 349-358. DOI: 10.1016/j.tree.2011.03.016 |
| [23] | Collins S L, Suding K N, Cleland E E, et al. Rank clocks and plant community dynamics[J]. Ecology, 2008, 89(12): 3534-3541. DOI: 10.1890/07-1646.1 |
| [24] | Gorelick R, Bertram S M, Killeen P R, et al. Normalized mutual entropy in biology:quantifying division of labor[J]. The American Naturalist, 2004, 164(5): 677-682. DOI: 10.1086/424968 |
| [25] | Shannon C E. A mathematical theory of communication[J]. Bell System Technical Journal, 1948, 27: 379-423, 623-656. DOI: 10.1002/bltj.1948.27.issue-3 |
| [26] | Lynam C P, Lilley M K S, Bastian T, et al. Have jellyfish in the Irish Sea benefited from climate change and overfishing?[J]. Global Change Biology, 2011, 17(2): 767-782. DOI: 10.1111/gcb.2010.17.issue-2 |
| [27] | Benton M J, Twitchett R J. How to kill(almost) all life:the end-Permian extinction event[J]. Trends in Ecology and Evolution, 2003, 18(3): 358-365. |
| [28] | Erwin D H. The end and the beginning:recoveries from mass extinctions[J]. Trends in Ecology and Evolution, 1998, 13(9): 344-349. DOI: 10.1016/S0169-5347(98)01436-0 |
| [29] | Magurran A E, Henderson P A. Explaining the excess of rare species in natural species abundance distributions[J]. Nature, 2003, 422(6933): 714-716. DOI: 10.1038/nature01547 |
| [30] | Yedid G, Ofria C A, Lenski R E. Selective press extinctions, but not random pulse extinctions, cause delayed ecological recovery in communities of digital organisms[J]. The American Naturalist, 2009, 173(4): E139-E154. DOI: 10.1086/597228 |
| [31] | Magurran A E, Henderson P A. Explaining the excess of rare species in natural species abundance distributions[J]. Nature, 2003, 422(6933): 714-716. DOI: 10.1038/nature01547 |
| [32] | Codron J, Botha-Brink J, Codron D, et al. Predator-prey interactions among permo-triassic terrestrial vertebrates as a deterministic factor influencing faunal collapse and turnover[J]. Journal of Evolutionary Biology, 2016, 30(1): 40-54. |
| [33] | Marshall C R. How stable are food webs during a mass extinction?[J]. Science, 2015, 350(6256): 38-39. DOI: 10.1126/science.aad2729 |