 2017, Vol. 53
2017, Vol. 53文章信息
- 张帅楠, 栾启福, 姜景民
- Zhang Shuainan, Luan Qifu, Jiang Jingmin
- 基于无损检测技术的湿地松生长及材性性状遗传变异分析
- Genetic Variation Analysis for Growth and Wood Properties of Slash Pine Based on The Non-Destructive Testing Technologies
- 林业科学, 2017, 53(6): 30-36.
- Scientia Silvae Sinicae, 2017, 53(6): 30-36.
- DOI: 10.11707/j.1001-7488.20170604
-
文章历史
- 收稿日期:2016-06-16
- 修回日期:2016-07-15
-
作者相关文章



湿地松(Pinus elliottii)是我国南方红壤低丘地区引种成功的速生用材、采脂树种,其成林早、生长快、树干通直、产脂量高(朱志淞等,1993),当前人工林面积达到300万hm2。由于我国湿地松人工林主要位于亚热带季风气候区,容易受到起伏不平的地形和夏季台风、冬季低温冰冻影响,在极端气候条件下弯折、倒伏风险较高(栾启福等,2008),因此针对我国造林地区特点,加强湿地松木材强度性状选育对于我国湿地松人工林健康持续发展具有积极意义。
木材基本密度(basic wood density,ρ)、弹性模量(modulus of elasticity,MOE)等性状是木材强度的重要测量指标,也是建筑材、纸浆材重要选择性状(Zobel et al., 1989)。然而受限于传统测量方法,在湿地松材性性状改良进程中,尤其是涉及到破坏性取样的弹性模量等力学性状每次测量的样品量较少(吴际友等,2000;徐有明等,2001;骆秀琴等,2003)。从统计角度而言其结果误差较大,这增大了遗传选择错误或遗漏的概率。因此,开发利用操作简单方便、破坏性小且结果可靠的无损评估技术,以大样本试验材料为基础,研究弹性模量等材性性状的遗传变异规律,对于提高遗传测定的选择精度与效率,促进遗传改良进程具有重大意义。近年来基于力学、声学的无损检测技术为大样本量估测活立木基本密度和弹性模量奠定了基础,逐渐成为国际上相关性状测量、选择的常规手段(White et al., 2007)。其中已成功应用于活立木弹性模量无损评估的是基于应力波速法的AV技术,它通过测量应力波在开始探头和结束探头之间的传播时间来测得应力波速v,该数值与立木强度具有显著的相关性,可以作为活立木强度的检测工具;Pilodyn是应用最为普遍的一种基于阻力的活立木密度无损快速检测仪器(Lenz et al., 2013;易敏等,2014;Chen et al., 2015)。
基于这2种简洁快速测量方法,本研究着重加强湿地松弹性模量遗传变异研究,在国内首次实现了2 000多株22年生湿地松半同胞家系全林每木基本密度、弹性模量的测量,同时按照传统方法测量了树高、胸径,并估测了其冠幅、枝下高和分枝大小等性状。通过无偏估计湿地松遗传群体主要物理力学性状、形态性状和生长量性状之间的遗传及其相关特性,为湿地松大群体活立木多性状综合评价及联合改良提供依据,这对于我国湿地松人工林健康持续发展具有积极意义。
1 材料与方法 1.1 试验林概况试验地位于浙江省杭州市余杭区长乐林场,30°27′ N,119°48′E,亚热带湿润季风性气候,年均温16.1 ℃,年均降水量1 399 mm,属低山缓坡,土壤以红壤为主。试验林营建于1994年春,为湿地松子代测定林,含有158个半同胞家系,试验布置采取完全随机区组设计,6株单行小区,6个重复,株行距2 m×3 m。根据培育规程,在林分10年生时对其进行隔株间伐,现保留单株共计2 500余株。
1.2 数据测定方法1) 生长性状测定:2015年12月,在林分停止生长期,采用测杆等标准测具对胸径、树高、枝下高、冠幅、分枝性状等主要生长性状进行全林每木调查,其中分枝大小和分枝角均按3级评测标准(1表示分枝较小或分枝角为0°~30°,3表示分枝较大或分枝角为60°~90°),单株材积按V=0.375H×DBH2估算(栾启福等,2011)。
2) 材性无损测定:利用Pilodyn(6J,PROCEQ,Switzerland)测定获得样木Pilodyn值(P)(朱景乐等,2008),P与木材基本密度(ρ)呈显著线性负相关关系,因此本文中将P作为基本密度相对值ρP(栾启福等,2011);利用活立木应力波速测量专用工具Hitman(Fibre-gen,Christchurch,New Zealand)对选择的样木进行测量获得波速v(Fibre-gen,2004;Carter et al., 2005),并利用弹性模量(MOE)、ρ和v之间的关系式MOE=ρ×v2(Ross et al., 1988)计算出弹性模量相对值MOEP(MOEP=1/P * v2)(Chen et al., 2015)。
1.3 统计分析方法1) 描述性数据分析利用R软件(v 3.1.3,CRAN,http://cran.r-project.org)中pastecs程序包的stat.desc()函数进行(林元震等,2014)。
2) 通过ASReml-R软件,利用其混合线性模型的限制性极大似然估计法(REML)对各性状的数据进行方差分析、遗传力估算和相关性分析(Butler et al., 2009;林元震等,2014)。该线性模型如下:
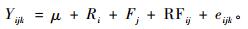
|
(1) |
式中:Yijk表示第ijk株树的性状观测值,μ表示该性状观察值的总体平均值,Ri表示重复效应,Fj表示家系效应,RFij表示重复和家系交互效应,eijk表示误差效应,其中Ri作为模型固定效应,Fj和RFij作为模型随机效应。
性状狭义遗传力估算公式为:
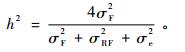
|
(2) |
式中:σF2、σRF2、σe2分别表示各性状的家系效应、重复和家系交互效应及标准误的方差组分。
性状遗传相关分析计算公式为:
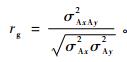
|
(3) |
式中:σAxAy2表示两性状间的加性协方差组分,σAx2和σAy2分别表示两性状的加性方差组分。
性状表型相关分析计算公式为:
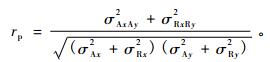
|
(4) |
式中:σRxRy2表示两性状间的残差协方差组分,σRx2和σRy2分别表示两性状的残差方差组分。
性状指标遗传增益计算公式(Namkoong,1981)为:

|
(5) |
式中:S、X和ha2分别代表每个性状指标的选择差、平均值和遗传力(入选率为1%)。
相关性状间间接选择有效性计算公式(White et al., 2007)为:
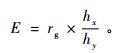
|
(6) |
式中: rg表示性状x、y之间的遗传相关关系,hx、hy分别表示性状x、y的遗传力的平方根,其中y是目标性状。
2 结果与分析 2.1 生长和材性性状各指标的表型变异分析2000余株湿地松的生长和材性性状的基本概况见表 1。22年生湿地松的胸径、树高、枝下高、材积和冠幅分别为20.45 cm,16.35 m,9.55 m,0.277 m3和3.26 m,枝下高约占整个树高的2/3左右,即湿地松冠层较小,单株树木建筑材质量和产量较高。其中胸径、树高、枝下高和冠幅变异系数(14.15%~26.99%)较小,材积变异系数达到了50%以上,说明材积观测值变异幅度大,具有较好的选择效果。弹性模量相对值MOEP的变异系数(31.60%)远大于基本密度相对值ρP(10.06%),这表明了与ρ相比,MOE可能具有更丰富的遗传变异性。
|
|
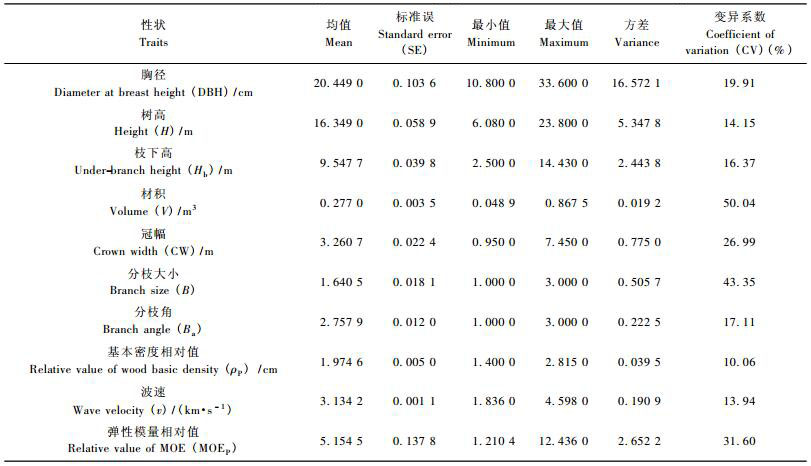
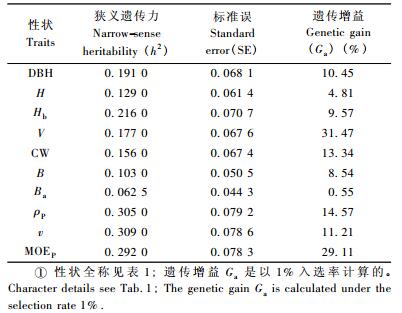
表 2显示了通过方差组分估算出的各性状狭义遗传力大小。材性性状指标的遗传力为0.292 0~0.309 0,明显高于生长性状的遗传力(0.062 5~0.216 0),即ρ和MOE受相对较高的遗传控制,而生长性状更易受环境影响,适宜的生长环境或栽培管理技术可以获得一定的生长增益。表 2还显示了以1%入选率对各性状进行直接选择时,各性状所能获得的遗传增益大小(其中,ρP的遗传增益计算时以ρP值最小的前1%个个体作为入选样本)。可以看出,材积(31.47%)和MOEP(29.11%)能获得较大增益,具有良好的选择效应;ρP的遗传增益(14.57%)较小,但是ρ的微小变化对最终木材产品的变化和效益仍是值得重视的(Zobel et al., 1972),因此湿地松ρ也具有较大的选择效益。
|
|
对湿地松生长和材性性状各指标进行遗传和表型相关分析(表 3)。就生长性状而言,胸径、材积、冠幅和分枝大小两两之间均呈显著的遗传正相关关系(R=0.440 0~0.995 0,P<0.05);树高与胸径(R=0.753 0 ± 0.134 0)、材积(R=0.865 0 ± 0.085 7) 之间呈极显著(P<0.001) 遗传正相关关系;枝下高仅与树高呈显著的遗传正相关性(R=0.436 0 ± 0.236 0,P<0.05)。分枝角除与分枝大小呈现极显著表型负相关关系(R=-0.147 0 ± 0.025 1,P<0.001) 外,与其他性状间均无显著表型或遗传相关关系;而胸径、树高、枝下高、材积、冠幅和分枝大小两两之间均具有极显著(P<0.001) 的表型正相关性。
|
|

同时,相关分析结果还显示材性和生长性状之间遗传相关不显著,但普遍存在着显著的表型正相关,显示材性和生长性状的遗传改良可同步进行,培育措施与生长和材性表现有关。对于材性性状无损测定指标,ρP与v之间遗传相关不显著,仅表型相关(R=-0.264 0 ± 0.025 1,P<0.001) 达到了显著性水平;MOEP和v之间呈极显著(P<0.001) 的遗传和表型正相关关系,相关系数分别达到0.947 0和0.948 0,以v对MOEP进行间接选择,其有效性达到了97.42%。MOEP和ρP之间呈显著(P<0.01) 遗传负相关(R=-0.447 0±0.154 0) 和极显著(P<0.001) 表型负相关(R=-0.538 0±0.019 1),即MOE和ρ之间显著正相关,但其与ρ的相关系数远小于与v的相关系数。
3 讨论弹性模量(MOE)是林木建筑用材最重要的力学性质(Raymond et al., 2004;Lasserre et al., 2007),然而受传统评估方法等限制,不同地区对湿地松MOE的改良均处于非优先地位。美洲湿地松人工林林地地势平坦,集约经营强度高,湿地松遗传改良主要集中在材积和抗锈病选择育种上(White et al., 2004),兼顾密度(ρ)等材性性状。国内对湿地松木材材性方面的相关研究也集中在生长性状,兼顾密度性状,对MOE等力学性状的遗传分析研究鲜有报道。单方面对生长性状的追求则被认为会严重影响木材的质量(Kumar,2004),不利于木材质量的提高。特别是我国南方湿地松人工林林地主要是低山丘陵,且集约化经营程度不高,容易受到起伏不平的地形和夏季台风、冬季低温冰冻影响,在极端气候条件下弯折、倒伏风险较高(栾启福等,2008)。对ρ、MOE等影响木材强度的性状的改良对于湿地松木材质量提高及其树种的健康持续发展具有积极意义。
本试验基于全林单株测定,采用无损评估方法,在国内首次实现了大规模湿地松活立木生长和材性的遗传变异分析。遗传力分析结果表明,湿地松ρ和MOE狭义遗传力约为0.30,受中等程度的遗传控制,与Chen等(2015)基于无损评估方法对5 000余株欧洲云杉(Picea abies)材性遗传分析结果较一致;而明显高于骆秀琴等(2003)基于常规检测方法得出的研究结果(湿地松弹性模量广义遗传力为0.108 2),这可能是由于在其研究中所取样木少(18个种源,共计90个单株)而导致的统计结果误差大造成的,而且本文中估算MOE的狭义遗传力标准误较低(SE=0.078 3),从侧面反映了大样本量下估算遗传参数的精确度和可靠性更高。其中,研究结果得出对MOE进行直接选择所能获得的遗传增益较大,这可能是由于此前树种改良工作中受研究条件限制没有对MOE进行直接选育,造成试验林内各家系或单株间MOE差异较大引起的。这表明急需加强湿地松MOE的遗传改良研究,加快湿地松材性选育工作进程,摒弃MOE表现不良的单株或家系,进而提高湿地松MOE或材性性状的整体水平,为湿地松建筑用材的选育奠定基础。
同时,了解生长与材性性状的遗传相关性有助于速生优质林木的选育,然而不同树种或研究方法对生长与材性性状的相关性研究结果不一致(Fries,1986;姜景民等,1996;Ukrainetz et al., 2008;刘青华等,2009),因此一直以来这都是育种工作者研究的重点。本文中相关性分析结果表明湿地松材性和生长性状之间遗传相关不显著,这与Fries(1986)和Wang等(1999)对小干松(Pinus contorta ssp. latifolia)以及吴际友等(2010)对湿地松生长和材性性状的相关性研究结果相同,可以实现二者的同步改良。同时,湿地松材性和生长性状之间还普遍存在着显著的表型正相关性,说明在特定的环境条件影响下,材性和生长性状之间能够呈现一定的正相关关系,这有利于二者的同步选择,即在实际生产上能够实现生长量大且材质优良湿地松家系或单株的选育。
相关性分析还发现应力波速(v)和MOE相对值(MOEP)之间呈高度的极显著遗传和表型正相关,以v对MOEP进行间接选择,其有效性达到了97.42%。这表明在比较同一测定群体MOE值相对大小时直接比较v值即可,即仅利用AV技术即能快速评估MOE相对值,这大大减少了同一群体内MOE性状评估与选择的工作量。然而,在此遗传变异分析研究过程中,所取样本量越大,研究所需的人力、物力就相对越多,因此,比较分析生长、材性等性状在不同取样规模下遗传参数估算的差异性,论证取样规模对其性状遗传参数的影响,进而确定各性状遗传参数评估所需的最适样本规模便成为了本研究进一步探讨的关键问题。
4 结论本文通过Pilodyn和应力波速技术等无损方法,以全林单株测定为基础,大规模评估了湿地松半同胞家系活立木的基本密度(ρ)和弹性模量(MOE)等材性性状,并通过遗传和相关分析,揭示了湿地松生长和材性的遗传变异及其相关关系。
1) 22年生湿地松的胸径、树高、枝下高、材积和冠幅等主要生长性状分别为20.45 cm、16.35 m、9.55 m、0.277 m3和3.26 m,枝下高约占整个树高的2/3左右,即湿地松冠层较小,有利于建筑材产量提高。材积观测值变异幅度大,具有较好的选择效果。与ρ相比,MOE具有更丰富的遗传变异性。
2) ρ和MOE受中等程度的遗传控制,其遗传力明显高于生长性状,这说明相较于材性性状,生长性状更易受环境影响。通过一定的选择强度,材积、ρ和MOE能获得较大增益,具有良好的选择效果。同时,本研究结果还显示材性和生长性状之间仅存在着显著的表型相关,能够实现两类性状同步遗传改良。
3) 应力波速(v)和MOE相对值(MOEP)之间呈高度的遗传和表型正相关关系,相关系数均达到了极显著水平,仅利用v即能快速评估群体内每木MOE相对大小,该方法比传统上仅用ρ评估木材MOE准确率高。
| [] |
姜景民, 孙海菁, 刘昭息. 1996. 火炬松纸浆材优良家系多性状选择. 林业科学研究, 9(5): 455–460.
( Jiang J M, Sun H Q, Liu Z X. 1996. Multitraits selection of loblolly pine families for pulpwood. Forest Research, 9(5): 455–460. [in Chinese] ) |
| [] |
林元震, 陈晓阳. 2014. R与ASReml-R统计分析教程. 北京, 中国林业出版社: 36-251.
( Lin Y Z, Chen X Y. 2014. R & ASReml-R statistical analysis tutorial. Beijing, China Forestry Publishing House: 36-251. [in Chinese] ) |
| [] |
刘青华, 金国庆, 张蕊, 等. 2009. 24年生马尾松生长、形质和木材基本密度的种源变异与种源区划. 林业科学, 45(10): 55–61.
( Liu Q H, Jin G Q, Zhang R, et al. 2009. Provenance variation in growth, stem-form and wood density of Masson pine at 24-year-old and the provenance division. Scientia Silvae Sinicae, 45(10): 55–61. DOI:10.11707/j.1001-7488.20091010 [in Chinese] ) |
| [] |
栾启福, 姜景民, 张建忠, 等. 2011. 火炬松×加勒比松F1代生长、树干通直度和基本密度遗传和配合力分析. 林业科学, 47(3): 178–183.
( Luan Q F, Jiang J M, Zhang J Z, et al. 2011. Estimation of heritability and combining ability for growth, stem-straightness and wood density of the F1 generation of Pinus taeda×P. caribaea. Scientia Silvae Sinicae, 47(3): 178–183. DOI:10.11707/j.1001-7488.20110327 [in Chinese] ) |
| [] |
栾启福, 卢萍, 肖复明, 等. 2008. 雨雪冰冻重灾区湿地松受害情况调查及其原因初步分析. 林业科学, 44(11): 50–54.
( Luan Q F, Lu P, Xiao F M, et al. 2008. Investigation on the damage of Pinus elliottii in the freezing rain and snow area and the analysis on the reason. Scientia Silvae Sinicae, 44(11): 50–54. DOI:10.3321/j.issn:1001-7488.2008.11.010 [in Chinese] ) |
| [] |
骆秀琴, 姜笑梅, 殷亚方, 等. 2003. 湿地松15个家系木材材性遗传变异及优良家系评估. 林业科学研究, 16(6): 694–699.
( Luo X Q, Jiang X M, Yin Y F, et al. 2003. Genetic variation and comprehensive assessment in wood properties of 15 families of Pinus elliottii. Forest Research, 16(6): 694–699. [in Chinese] ) |
| [] |
吴际友, 李志辉, 龙应忠, 等. 2010. 湿地松全同胞家系主要经济性状的遗传变异与选择研究. 中南林业科技大学学报, 30(8): 1–4.
( Wu J Y, Li Z H, Long Y Z, et al. 2010. Genetic variation and selection of main economic characteristics of slash pine full-sib family. Journal of Central South University of Forestry & Technology, 30(8): 1–4. [in Chinese] ) |
| [] |
吴际友, 龙应忠, 余格非, 等. 2000. 湿地松半同胞家系主要经济性状的遗传分析及联合选择. 林业科学, 36(1): 56–61.
( Wu J Y, Long Y Z, Yu G F, et al. 2000. Genetics analysis and combined selection of main economic characters of half-sib families for slash pine. Scientia Silvae Sinicae, 36(1): 56–61. [in Chinese] ) |
| [] |
徐有明, 鲍春红, 周志翔, 等. 2001. 湿地松种源生长量、材性的变异与优良种源综合选择. 东北林业大学学报, 29(5): 18–21.
( Xu Y M, Bao C H, Zhou Z X, et al. 2001. Variation analyses of tree growth and wood properties among slash pine provenances and comprehensive selection of superior provenance. Journal of Northeast Forestry University, 29(5): 18–21. [in Chinese] ) |
| [] |
易敏, 赖猛, 孙晓梅, 等. 2014. 无损检测在日本落叶松材性育种中的应用前景探讨. 林业科学, 50(11): 96–103.
( Yi M, Lai M, Sun X M, et al. 2014. Application prospect on applying nondestructive testing technology in wood property breeding of Larix kaempferi. Scientia Silvae Sinicae, 50(11): 96–103. [in Chinese] ) |
| [] |
朱景乐, 王军辉, 张守攻, 等. 2008. 毛白杨材性指标预测及选择. 林业科学, 44(7): 23–28.
( Zhu J L, Wang J H, Zhang S G, et al. 2008. Wood property estimation and selection of Populus tomentosa. Scientia Silvae Sinicae, 44(7): 23–28. DOI:10.11707/j.1001-7488.20080705 [in Chinese] ) |
| [] |
朱志淞, 丁衍畴. 1993. 湿地松. 广州, 广东科技出版社: 62-298.
( Zhu Z S, Ding Y C. 1993. Slash pine. Guangzhou, Guangdong Science & Technology Press: 62-298. [in Chinese] ) |
| [] | Butler D G, Cullis B R, Gilmour A R, et al. 2009. ASReml-R reference manual. Queensland Department of Primary Industries. |
| [] | Carter P, Briggs D, Ross R J, et al. 2005. Acoustic testing to enhance Western forest values and meet customer wood quality needs//Harrington C A, Schoenholtz S H. Productivity of Western forests: a forest products focus. General technical report PNW-GTR-642. Portland: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research, 121-129. |
| [] | Chen Z Q, Karlsson B, Lundqvist S O, et al. 2015. Estimating solid wood properties using Pilodyn and acoustic velocity on standing trees of Norway spruce. Annals of Forest Science, 72(4): 1–10. |
| [] | Fibre-gen. 2004. Director ST300-operating manual. http://www.fibre-gen.com/documents/ST300OperatingManual.pdf. |
| [] | Fries A. 1986. Volume growth and wood density of plus tree progenies of Pinus contorta in two Swedish field trials. Scand J For Res, 1(1/4): 403–419. |
| [] | Kumar S. 2004. Genetic parameter estimates for wood stiffness, strength, internal checking, and resin bleeding for radiata pine. Canadian Journal of Forest Research, 34(12): 2601–2610. DOI:10.1139/x04-128 |
| [] | Lasserre J P, Mason E G, Watt M S. 2007. Assessing corewood acoustic velocity and modulus of elasticity with two impact based instruments in 11-year-old trees from a clonal-spacing experiment of Pinus radiata D. Don. Forest Ecology and Management, 239: 217–221. DOI:10.1016/j.foreco.2006.12.009 |
| [] | Lenz P, Auty D, Achim A, et al. 2013. Genetic improvement of white spruce mechanical wood traits—early screening by means of acoustic velocity. Forests, 4(3): 575–594. DOI:10.3390/f4030575 |
| [] | Namkoong G. 1981. Introduction to quantitative genetics in forestry. London, Castle House Publications. |
| [] | Raymond C A, Kube P D, Pinkard L, et al. 2004. Evaluation of non-destructive methods of measuring growth stress in Eucalyptus globulus: relationships between strain, wood properties and stress. Forest Ecology and Management, 190(2): 187–200. |
| [] | Ross R J, Pellerin R F. 1988. NDE of wood-based composites with longitudinal stress wave. Forest Products Journal, 38(5): 39–45. |
| [] | Ukrainetz N K, Kang K Y, Aitken S N, et al. 2008. Heritability and phenotypic and genetic correlations of coastal Douglas-fir (Pseudotsuga menziesii) wood quality traits. Canadian Journal of Forest Research, 38(6): 1536–1546. DOI:10.1139/X07-234 |
| [] | Wang T, Aitken S N, Rozenberg P, et al. 1999. Selection for height growth and Pilodyn pin penetration in lodgepole pine: effects on growth traits, wood properties and their relationships. Can J For Res, 29: 434–445. DOI:10.1139/x99-012 |
| [] | White T L, Adams W T, Neale D B.2007. Forest genetics. CABI, Wallingford. |
| [] | White T L, Byram T D. 2004. Slash pine tree improvement//Dickens E D, Barnett J P, Hubbard W G, et al. Proceedings of the slash pine symposium. Slash pine: still growing and growing! SRS-76. Asheville, NC: U.S. Department of Agriculture, Forest Service, Southern Research Station, 7-19. |
| [] | Zobel B J, Kellison R C, Matthias M F, et al. 1972. Wood density of the southern pines. North Carolina Agricultural Experiment Station.Tech Bul 208:56. |
| [] | Zobel B J, Van Buijtenen J P. 1989. Wood variation: its causes and control. Springer Science & Business Media. |