 2016, Vol. 52
2016, Vol. 52文章信息
- 刘自强, 余新晓, 贾国栋, 贾剑波, 娄源海, 张坤
- Liu Ziqiang, Yu Xinxiao, Jia Guodong, Jia Jianbo, Lou Yuanhai, Zhang Kun
- 北京山区侧柏和栓皮栎的水分利用特征
- Water Use Characteristics of Platycladus orientalis and Quercus variabilis in Beijing Mountain Area
- 林业科学, 2016, 52(9): 22-30
- Scientia Silvae Sinicae, 2016, 52(9): 22-30.
- DOI: 10.11707/j.1001-7488.20160903
-
文章历史
- 收稿日期:2015-10-08
- 修回日期:2015-11-17
-
作者相关文章



2. 邢台县水务局 邢台 054001
2. Water Affairs Bureau of Xingtai County, Hebei Xingtai 054001
树木生态学特性和生理特征受多种因素制约,水分则是对树木生长发育影响最大的生态因子之一(张文辉,1998)。不同生活型和地理分布的树木采取不同的水分利用策略适应生境(陈洪松等,2013)。利用传统的物理方法研究树木对水分利用存在较大困难(巩国丽等,2011),而稳定氢氧同位素技术作为一种新的研究手段已广泛应用于生态学领域的树木水分来源研究中(Zhang et al.,2010)。除一些盐生植物外,树木根系对水分的吸收以及水分在木质部运输的过程不会出现同位素分馏现象(Ellsworth,2007; 石辉等,2003)。因此,对比分析树木枝条水分与各水源的同位素组成,参考土壤含水量等信息可推测树木的主要水分来源,再结合多元线性模型可进一步量化各水源对树木水分的贡献比例(聂云鹏等,2010; 段德玉等,2007)。目前,常利用稳定氢氧同位素技术分析植物水分来源,并判断植物对不同水源利用比例。沙漠地区的乔木存在多种水分利用模式(戴岳等,2014);不同季节对不同水源的利用存在转换(邢星等,2014; 朱雅娟等,2010);在准噶尔盆地发现多枝柽柳(Tamarix ramosissima)、白刺(Nitraria sibirica)和红砂(Reaumuria soongorica)3种树木的水分来源有明显的季节波动特征(周海等,2013);季节性干旱热带林区,常绿树种通过加速生长的深根系吸收深层土壤水分(Hasselquist et al.,2010)。水分较为充足的区域,植物更倾向于利用浅层土壤水(周雅聃,2011)。邓文平等(2014)利用氢氧同位素分析了雨季北京山区3种典型树种的水分来源。还有些研究结合树木的生理指标阐释树木对异质性生境的适应机制(Yang et al.,2011; Penuelas et al.,2011; 孙守家等,2011)。多数学者利用稳定氢氧同位素研究不同生活型植物的水分来源及其对各层土壤水分的利用率,较少学者利用稳定氢氧同位素技术,结合茎干液流计(TDP)测算不同生活型树木对各层土壤水分的利用量。
北京山区是华北平原和北京市区的重要生态屏障,但其处于季节性干旱的生态脆弱地带,水源短缺,气候易旱,年降水分布不均,土层瘠薄,了解其典型树种在干旱生境下正常生长发育的水分利用机制具有重要意义。本文以北京山区分布较广的侧柏(Platycladus orientalis)和栓皮栎(Quercus variabilis)典型人工树种为研究对象,对林地土壤水分和树木水分的氢氧同位素组成进行分析,再结合树干液流计(TDP)测算树木液流通量,探讨北京山区典型树木对不同土层的土壤水分利用率和利用量,以期为人工造林树种选择、配置及其后期管理提供科学参考。
1 研究区概况研究区位于鹫峰国家森林公园——首都圈森林生态系统定位研究站。海拔约450 m,40°03′N,116°05′E,属于华北暖温带半湿润半干旱大陆性季风气候区。多年平均降水量630 m,但全年降水分布不均,主要集中在6—9月,该时间段的降水量可达全年降水量的80%;年蒸发量高达1 800~2 000 mm。该区森林植被基本以1958年以后营造的人工林为主,是乔木灌木的混交林,全园森林覆盖率85%左右,土壤类型为淋溶褐土,土层较薄,土壤表层腐殖质含量较高,较深层土壤基本为石砾(邓文平,2013)。试验地及标准株基本情况如图 1。

|
图 1 取样点分布 Fig.1 Location of sampling site within the topographic map of the studied area |
|
|
本研究的样品采集时间为2012年旱季04月17日和雨季07月20日。
在2块样地内分别选择3株无人工干扰的、长势良好且相似的,胸径、树高和形数在林分平均水平的标准木,采集其已栓化的、成熟的枝条(直径约3 cm),每株采集3个样品。在标准植株附近,分别选取3个的采样点利用土钻采集土壤样品。土壤表层40 cm以上每间隔10 cm采集土壤样品,40 cm以下每间隔20 cm采集土壤样品,每层采集3个样品。收集样点附近的泉水水代表样点的地下水,泉水采集点处于金山泉和妙灵山泉,每处泉水设置3个采集点,每个采集点采集3次。为防止所采集样品水分蒸发导致同位素分馏,将其迅速放入瓶中,用Parafilm膜密封、冷藏。
2.2 同位素样品的真空抽提釆用真空抽提装罝来抽提枝条和土壤中的水分(Jia et al.,2012),装置包含5个独立的抽提单元,并都连接于玻璃主管道。将装有枝条或土壤的10 mL样品瓶放入底部装有小团棉花的抽提管,并在样品瓶的上方塞一棉花,防止抽提过程土壤因过热喷入上方玻璃管壁。随后将连接在玻璃主管道的抽提管浸没在装有液氮的杜瓦瓶冷凝10~15 min。冷凝完成后,打开真空泵,对管路内的空气进行真空抽提,当真空度达到10 m Torr以下时移去抽提管上的杜瓦瓶,紧接着给其套上己恒温至95 ℃的加热套,并将装有液氮的杜瓦瓶浸过收集管的底部。收集的过程中不断的添加液氮保证杜瓦瓶中液氮始终处于恒定的高度,最后将收集管用Parafilm膜密封、冷藏。
2.3 样品同位素值测定除去所采枝条的表皮和皮层,然后采用低温真空冷凝法抽提枝条和土壤水分。对含水量较低的土壤样品,抽提出来的水分不足以满足测试所需的最低用量时,需要重复抽提该样品。随后用 LGR DLI-100液态水同位素分析仪测定抽出水样中的同位素δD,δ18O值,其δD,δ18O的测定精度分别达0.3‰和0.1‰。测得水样中的氢氧同位素含量为与“标准平均大洋水(SMOW)”的千分差,表示为:
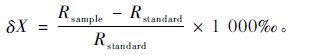
|
式中,δX 为δD或δ18O。Rsample和Rstandard分别表示样品和SMOW中的D或18O的浓度。
当树木具有多个不同水分来源时,树木茎干水分的同位素值为多个不同水源同位素值的混合值(Jackson et al.,1999),可以根据同位素质量守恒原理,采用多元线性分析方法,利用Iso-Source模型计算各水源对树木茎干水分的贡献率。计算时,将各潜在水源的D和18O同位素同时输入Iso-Source模型;当2个潜在水源的同位素值相近或相同时,将2个水源合并为1个。计算公式为:
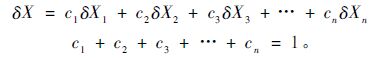
|
(2) |
式中,δX为树木茎干木质部水分δD或δ18O值,‰;c1,c2,c3…cn分别表示各水源对树木所利用水分的贡献率;X1,X2,X3…Xn分别为各水源水分的δD或δ18O值。
2.4 茎干液流测定选取热扩散原理(插针式)茎流计测定树木茎干液流速率。先用砂纸将所选侧柏和栓皮栎树木茎干打磨光滑,再在打磨区涂抹G4油脂。将探头安装在被测区域后用绝缘、防辐射材料包裹、密封,防止雨水进入。具体测定原理及具体安装方法见黄磊等(2011)。安装好后,将数据传输线与数据采集器连接,接通电源,设置相关参数,定期维护,用计算机进行数据采集。
本试验设置数据采集间隔时间为15 min,选取04月17日和07月20日全天的侧柏和栓皮栎的茎干液流监测数据。树木的液流通量计算公式为:
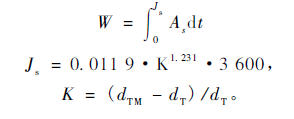
|
式中,W为液流通量(cm3);Js为液流密度(cm3·cm-2h-1);As为边材面积(cm2);t为对应液流密度的时间;K为无量纲的常数;dTM为液流速率为零时的探针最大温差值;dT为当时测定的温差值。
3 结果与分析 3.1 侧柏、栓皮栎林地土壤含水量特征不同季节2块林地土壤含水量不同(图 2);在旱季和雨季,栓皮栎林地土壤含水量均比侧柏林地土壤含水量高,这可能由于侧柏为常绿树种,林下枯枝落叶少,且难以分解,而栓皮栎阔叶林有大量的枯枝落叶覆盖在土壤表层,使其土壤含水量较高。在侧柏林地,雨季受降雨影响,其土壤含水量相对于旱季高148.84%;旱季侧柏林地表层0~10 cm土壤受蒸发影响,其含水量较10~30 cm土壤低70.39%;40~100 cm土壤含水量几乎相同,平均值为 1.72%;雨季侧柏林地0~10 cm土壤含水量较高,为6.14%;10~20 cm及以下土壤含水量减少,这可能与侧柏对该层土壤水分吸收及土壤理化性质有关;50~100 cm土壤含水量在5.21%左右变动。在栓皮栎林地,雨季土壤含水量比旱季土壤含水量大185.81%;旱季栓皮栎林地表层土壤蒸发强烈,0~20 cm土壤含水量为3.04%,20~60 cm土壤含水量变化不大,平均值为 6.43%;雨季时期0~30 cm土壤,随着土壤深度的增加,土壤含水量减小,这可能是由于降水入渗到深层土壤的水分减少造成的;30~60 cm土壤含水量变化不大,平均值16.19%。

|
图 2 侧柏、栓皮栎林地土壤剖面含水量特征 Fig.2 The variation of soil water in P. orientalis and Q. variabilis forests A. 侧柏林地P. orientalis B. 栓皮栎林地Q. variabilis |
对比树木茎干与各层土壤水的δD值,可知不同季节侧柏和栓皮栎茎干的δD值均不同,说明不同树木在不同季节水分来源有差异(图 3);旱季蒸发量比雨季蒸发量大,导致2个树种在旱季茎干水分δD值都比雨季大;在旱季时期,栓皮栎茎干的δD值较侧柏茎干的δD值大,可能由于栓皮栎蒸发更强烈,导致重同位素富集。

|
图 3 树木茎干水分的同位素特征与潜在水源的关系 Fig.3 The relation of tree stem water isotope characteristics and potential of water A. 侧柏(4 月17 号) In April 17th of P. orientalis; B. 侧柏(7 月20 号) In July 20th of P. orientalis; C. 栓皮栎(4 月17 号) In April 17th of Q. variabilis; D. 栓皮栎(7 月20 号) In April 17th of Q. variabilis |
在旱季,侧柏和栓皮栎林地土壤随深度的增加,土壤水分中的δD值减小;侧柏茎干水的δD值与土壤20~30 cm处有1个交点,且与土壤70~80 cm水的δD值接近,结合土壤含水量变化可知该层土壤水分变化剧烈,说明侧柏吸收利用了这几层土壤水分;侧柏茎干水的δD值与土壤70~80 cm水的δD值相近,说明侧柏可能吸收利用了该层土壤水分;栓皮栎茎干水的δD值与土壤0~20 cm水的δD值相交,根据该层土壤含水量变化,可以推测栓皮栎吸收利用这层土壤水分。
在雨季侧柏茎干水的δD值与土壤水的δD值在0~10 cm及20~30 cm处相交,且侧柏茎干水的δD值与地下水的δD值接近,这可能是侧柏直接利用这3层水分所致。栓皮栎茎干水的δD值与土壤的δD值在0~10 cm及20~30 cm处相交,同时栓皮栎茎干水的δD值与70~80 cm土壤水的δD值相近,说明栓皮栎可能利用这3层土壤水分。
3.3 侧柏、栓皮栎的茎干液流通量不同树种在不同时期的茎干液流通量不同(图 4)。2个树种在旱雨季时期茎干液流通量的排序为:旱季侧柏(5.87 kg)<旱季栓皮栎(9.42 kg)<雨季侧柏(13.42 kg)<雨季栓皮栎(18.26 kg)。在相同时期,栓皮栎的液流通量大于侧柏;在旱季栓皮栎的液流通量比侧柏的液流通量高60.48%;在雨季栓皮栎的液流通量比侧柏的液流通量高36.07%,这是由于阔叶栓皮栎比针叶侧柏蒸发强烈造成的。2个树种在雨季时期的液流通量相对于旱季时期的液流通量都增加,侧柏增加128.62%,栓皮栎增加89.67%,结合上述土壤含水量分析,旱季时期的液流通量较低,这可能是树种为了适应干旱环境、较少水分蒸发而采取的保护性措施(王艳兵,2013),而雨季时期的土壤含水量大,处于生长季的树木能获得较为充沛的水分供给,导致雨季时期的液流通量大。

|
图 4 侧柏、栓皮栎的茎干液流通量 Fig.4 The stem sap flow flux of P. orientalis and Q. variabilis |
因上述不同树种在2个时期的土壤含水量及同位素特征不同,所以吸收利用不同的水源导致2个树种在不同时期对各水源的利用率和利用量均不相同。Iso-source软件分析结果表明(表 2),侧柏在旱季时期主要利用地下水,而对表层0~20 cm土壤相对利用较少,对地下水的利用率为51.4%,日平均利用量为3.02 kg;对表层0~20 cm土壤利用率为17.3%,日平均利用量为1.01 kg。雨季的侧柏对表层0~20 cm土壤水分的利用率相对旱季增加197.11%,日平均利用量为6.90 kg;雨季时期的侧柏对地下水的利用率为旱季时期的53.89%,但日平均利用量相对旱季有所增加,为3.72 kg。栓皮栎在旱季时期主要利用表层0~20 cm的土壤水分,利用率为53.5%,日平均利用量为5.14 kg;对土壤深层的水分利用较少,深层60~80 cm土壤对其贡献率为20.2%,贡献量为表层0~20 cm的37.69%。雨季的栓皮栎对表层0~20 cm土壤的利用率与旱季时期相差不大,为51.8%,但雨季时期对表层0~20 cm土壤的日平均利用量比旱季高79.40%,为9.46 kg。雨季时期的栓皮栎利用比60~80 cm土壤更为深层的地下水,利用率为38.1%,日平均利用量为6.96 kg。
|
|
林地土壤水分受蒸发和降水的双重影响,处于不断变化的状态(王海燕等,2013)。通过分析不同林地各层土壤水分含量和水同位素特征可知,2块林地由于植被类型以及土壤本身性质不同,土壤水分含量和水同位素特征也不相同。与侧柏林地相比,栓皮栎林地的地表被一层厚厚的枯落物覆盖,这使得土壤的蒸发量减弱、含水量高;也由于同一时期侧柏林地土壤的蒸发量比栓皮栎林地大,其重同位素出现富集,水同位素特征值较大。徐庆等(2007)研究四川卧龙山亚高山暗针叶林地区的各层土壤剖面水分氢氧稳定同位素特征值的变化规律及其水分含量的信息,认为土壤表层水的δD值受蒸发的直接影响,并且土壤水分同位素特征值显示出垂直变化。贾国栋(2013)研究北京山区的氢氧同位素值的变化规律,认为上层土壤受环境因子影响较大,δ18O和δD随土壤深度的增加而减小,深层土壤及浅层地下水的同位素特征值受环境因子的影响很小,δ18O和δD能保持稳定。旱季时期的侧柏林地和栓皮栎林地的土壤含水量低,水同位素特征值受水分蒸发影响较大,尤其是表层0~20 cm的土壤水同位素特征值,分馏效应明显;20~80 cm土壤水同位素特征值随着深度的加深而变小。雨季侧柏林地和栓皮栎林地的土壤含水量相对充足,随着土壤深度的增加,土壤水分同位素特征值显示出垂直变化;80 cm以下未受到季节影响的土壤水分同位素特征值与浅地下水相近。
不同季节时期的不同树种,其水分利用策略不同。侧柏在旱季时期主要利用深层土壤水分,在雨季利用表层0~20 cm的土壤水分;而栓皮栎在旱季的水分利用率贡献最大的还是集中在表层0~20 cm的土壤,在雨季的水分利用率贡献最大的还是集中在表层0~20 cm的土壤水和地下深层水分。侧柏和栓皮栎这2种植物的水分利用模式不同,与其根系的分布和根深有关,因为根系是植物吸收水分的主要方式之一,因此,根系的分布和根深对植物吸收利用水分有很大的影响(Lin et al.,1996)。土壤水分情况受不同植被根系分布特征影响,侧柏具有丰富的表层根系,约50%的细根系分布在0~20 cm处,但主根系不发达;而栓皮栎根系在各层土壤分布较为均匀,并具有较强的主根系(贾国栋,2013; 蔡鲁等,2015),侧柏在旱季时期吸收深层土壤水分维持正常的生理活动以适应干旱环境,但丰富的表层根系对降水反应敏感,使其在雨季集中利用表层土壤水分。结合2个树种在2个时期的液流通量分析,相对于旱季,雨季的土壤水势升高,土壤对水的吸持能力减弱,导致根系吸收水分的阻力变小,相似的大气条件下,土壤-植物-大气的水势梯度增加,增加了水分流动的驱动力,提升了茎干液流速率(冯永健等,2010),导致雨季的侧柏和栓皮栎液流通量比旱季时期高109.15%,这就出现了2个树种在旱季对某一层水分的利用率比雨季时期高,但其在旱季时期对某一土层水分的利用量很可能比雨季时期低的现象。无论旱季或雨季,阔叶树种栓皮栎的蒸腾耗水量都要大于侧柏,史宇(2011)和赵勇等(2010)也得出干旱半干旱区的栓皮栎蒸腾耗水量要大于侧柏,并且栓皮栎在雨季的蒸腾耗水量最大。对于旱季的栓皮栎来说,表层土壤水分基本能够满足它的蒸腾消耗,而且吸收表层土壤水分运输距离短,消耗的能量少,因此,更倾向于吸收利用表层土壤水分;雨季的栓皮栎为了维持较高的蒸腾强度,利用发达的主根系转而吸收利用深层土壤这一稳定的水源,说明栓皮栎对土壤深层水分可能有更强的依赖性。这2个树种对水分利用策略的不同可能是植物对环境长期适应的结果。
5 结论利用Iso-source多源线性混合模型,再结合侧柏和栓皮栎液流通量分析,得出2个树种对各潜在的利用特征。2个树种在旱雨季茎干液流通量的排序为:旱季的侧柏(5.87 kg)<旱季的栓皮栎(9.42 kg)<雨季的侧柏(13.42 kg)<雨季的栓皮栎(18.26 kg)。侧柏在旱季时期主要利用深层60~80 cm的土壤水分和地下水,利用率分别为19.8%和51.4%,日平均利用量为1.16和3.02 kg;在雨季时期主要利用表层0~20 cm的土壤水分和地下水,利用率分别为51.4%和27.7%,日平均利用量为6.90和3.72 kg。从侧柏的水分利用特征可知,旱季时期对地下水的利用率比雨季高46.10%,但利用量却与雨季时期相似。栓皮栎在旱季时期的水分利用率贡献最大的还是集中在表层0~20 cm的土壤水分和深层60~80 cm的土壤水分,利用率分别为53.5%和20.2%,日平均利用量为5.04和1.90 kg;在雨季时期的水分利用率贡献最大的还是集中在表层0~20 cm的土壤水和地下浅层水分,利用率分别为51.8%和38.1%,日平均利用量为9.46和6.96 kg。从栓皮栎的水分利用特征看出,2个时期对表层0~20 cm的土壤水分利用率相似,但雨季时期的利用量将近旱季时期的2倍。从侧柏和栓皮栎的水分利用特征可知,2个树种对各水源的利用深度相似,在造林树种选择时,因避免混交产生竞争。在季节性干旱的北京山区,水分匮缺,在造林时应优先选择蒸腾耗水较小的侧柏树种。
| [1] |
蔡鲁, 朱婉芮, 王华田, 等. 2015. 鲁中南山地6个造林树种根系形态的比较. 中国水土保持科学 , 13 (2) : 83–91.
( Cai L, Zhu W B, Wang H T, et al.2015. Root morphology of six tree species in mountain area of middle south Shandong. Science of soil and Water Conservation , 13 (2) : 83–91. [in Chinese] ) (  0) 0)
|
| [2] |
陈洪松, 聂云鹏, 王克林. 2013. 岩溶山区水分时空异质性及植物适应机理研究进展. 生态学报 , 33 (2) : 317–326.
( Chen H S, Nie Y P, Wang K L.2013. Spatio-temporal heterogeneity of water and plant adaptation mechanisms in karst regions:a review. Acta Ecologica Sinica , 33 (2) : 317–326. DOI:10.5846/stxb [in Chinese] ) ( 0)
|
| [3] |
戴岳, 郑新军, 唐立松, 等. 2014. 古尔班通古特沙漠南缘梭梭水分利用动态. 植物生态学报 , 38 (11) : 1214–1225.
( Dai Y, Zheng X J, Tang L S, et al.2014. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Chinese Journal of Plant Ecology , 38 (11) : 1214–1225. DOI:10.3724/SP.J.1258.2014.00117 [in Chinese] ) ( 0)
|
| [4] |
邓文平, 余新晓, 贾国栋, 等. 2013. 北京西山鹫峰地区氢氧稳定同位素特征分析. 水科学进展 , 24 (5) : 642–650.
( Deng W P, Yu X X, Jia G D, et al.2013. Analysis of hydrogen and oxygen stable isotopic characteristics of Beijing Xishan Jiufeng area. Advances in Water Science , 24 (5) : 642–650. [in Chinese] ) ( 0)
|
| [5] |
邓文平, 余新晓, 贾国栋, 等. 2014. 雨季北京山区3种典型植物的水分来源. 干旱区研究 , 31 (4) : 649–657.
( Deng W P, Yu X X, Jia G D, et al.2014. Water sources of 3 typical plants in Beijing mountain area during rainy season. Arid Zone Research , 31 (4) : 649–657. [in Chinese] ) ( 0)
|
| [6] |
段德玉, 欧阳华. 2007. 稳定氢氧同位素在定量区分植物水分利用来源中的应用. 生态环境 , 16 (2) : 655–660.
( Duan D Y, Ou Y H.2007. Application of stable hydrogen and oxygen isotope in analyzing plant water use sources. Ecology Environment , 16 (2) : 655–660. [in Chinese] ) ( 0)
|
| [7] |
冯永健, 马长明, 王彦辉, 等. 2010. 华北落叶松人工林蒸腾特征及其与土壤水势的关系. 中国水土保持科学 , 8 (1) : 93–98.
( Feng Y J, Ma C M, Wang Y H, et al.2010. Relationship between the characteristic of transpiration of Larix principi-rupprechtii fore stand soil water potential. Science of Soil and Water Conservation , 8 (1) : 93–98. [in Chinese] ) ( 0)
|
| [8] |
巩国丽, 陈辉, 段德玉. 2011. 利用稳定氢氧同位素定量区分白刺水分来源的方法比较. 生态学报 , 31 (24) : 7533–7541.
( Gong G L, Chen H, Duan D Y.2011. Comparison of the methods using stable hydrogen and oxygen isotope to distinguish the water source of Nitraria tangutorum. Acta Ecologica Sinica , 31 (24) : 7533–7541. [in Chinese] ) ( 0)
|
| [9] |
黄磊, 张志山. 2011. 荒漠人工植被区柠条和油蒿茎干液流动态研究. 中国沙漠 , 31 (3) : 683–688.
( Huang L, Zhang Z S.2011. The desert vegetation region Artemisia ordosica and Caragana microphylla stem sap flow dynamics. Journal of Desert Research , 31 (3) : 683–688. [in Chinese] ) ( 0)
|
| [10] |
贾国栋.2013.基于稳定氢氧同位素技术的植被-土壤系统水分运动机制研究.北京:北京林业大学博士学位论文. ( Jia G D.2013.Stable hydrogen and oxygen isotope technology of vegetation-study on soil water movement mechanism based on the system.Beijing:PhD thesis of Beijing Forestry University.[in Chinese][in Chinese]) |
| [11] |
聂云鹏, 陈洪松, 王克林. 2010. 土层浅薄地区植物水分来源研究方法. 应用生态学报 , 21 (9) : 2427–2433.
( Nie Y P, Chen H S, Wang K L.2010. Method for the study of plant water source in shallow soil. Chinese Journal of Applied Ecology , 21 (9) : 2427–2433. [in Chinese] ) ( 0)
|
| [12] |
石辉, 刘世荣, 赵晓广. 2003. 稳定性氢氧同位素在水分循环中的应用. 水土保持学报 , 17 (2) : 163–166.
( Shi H, Liu S R, Zhao X G.2003. Application of stable hydrogen and oxygen isotope in water circulating. Jourunal of Soil and Water Conservation , 17 (2) : 163–166. [in Chinese] ) ( 0)
|
| [13] |
史宇.2011.北京山区主要优势种森林生态系统的生态水文过程分析.北京:北京林业大学博士学位论文. ( Shi Y,2011.Eco-hydrological process analysis on forest ecosystems of major dominant species in Beijing mountainous area.Beijing:PhD thesis of Beijing Forestry University.[in Chinese][in Chinese]) |
| [14] |
孙守家, 孟平, 张劲松, 等. 2011. 利用氘同位素研究太行山南麓枣树水分利用的季节性变化. 林业科学 , 47 (5) : 46–53.
( Sun S J, Meng P, Zhang J S, et al.2011. Seasonal variation in water use of jujuba in the south aspect of taihang mountains with deuterium isotope signature. Scientia Silvae Sinicae , 47 (5) : 46–53. [in Chinese] ) ( 0)
|
| [15] |
邢星, 陈辉, 朱建佳, 等. 2014. 柴达木盆地诺木洪地区5种优势荒漠植物水分来源. 生态学报 , 34 (21) : 6277–6286.
( Xing X, Chen H, Zhu J J, et al.2014. Water sources of five dominant desert plant species in Nuomuhong area of Qaidam Basin. Acta Ecologica Sinica , 34 (21) : 6277–6286. [in Chinese] ) ( 0)
|
| [16] |
徐庆, 刘世荣, 安树青, 等. 2007. 四川卧龙亚高山暗针叶林土壤水的氢稳定同位素特征. 林业科学 , 43 (1) : 8–14.
( Xu Q, Liu S R, An S Q, et al.2007. Characteristics of hydrogen stable isotope in soil water of subalpine dark coniferous forest in Wolong,Sichuan Province. Scientia Silvae Sinicae , 43 (1) : 8–14. [in Chinese] ) ( 0)
|
| [17] |
王海燕, 刘廷玺, 王力, 等. 2013. 科尔沁沙地坨甸交错区土壤水分的空间变异规律. 干旱区研究 , 30 (3) : 438–443.
( Wang H Y, Liu T X, Wang L, et al.2013. Spatial variation of soil moisture content in the dune-meadow ecotone in the Horqin sand land. Arid Zone Research , 30 (3) : 438–443. [in Chinese] ) ( 0)
|
| [18] |
王艳兵.2013.六盘山半干旱区华北落叶松树干径向变化及对环境因子的响应.内蒙古:内蒙古农业大学硕士学位论文. ( Wang Y B.2013.Six Panshan radial variation in semi arid area of North China larch stem and the response to environmental factors.Inner Mongolia:MS Thesis of Agricultural University.[in Chinese][in Chinese]) |
| [19] |
张文辉. 1998. 裂叶沙参种群生态学研究. 哈尔滨: 东北林业大学出版社 .
( Zhang W H. 1998. lobophylla population ecology. Harbin: Northeast Forestry University Press . [in Chinese] ) |
| [20] |
赵勇, 陈桢, 樊巍, 等. 2010. 太行山低山丘陵区7种典型植物水分利用特征,中国水土保持科学,8(5):61-66. 中国水土保持科学 , 8 (5) : 61–66.
( ZhaoY, Chen Z, Fan W, et al.2010. Water utilization characteristics of 7 typical plant species in hilly region of Taihang Mountains. Science of Soil and Water Conservation , 8 (5) : 61–66. [in Chinese] ) ( 0)
|
| [21] |
周海, 郑新军, 唐立松, 等. 2013. 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同. 植物生态学报 , 37 (7) : 665–673.
( Zhou H, Zhen X J, Tang L S, et al.2013. Differences and similarities between water sources of Tamarix ramosissima,Nitraria sibirica and Reaumuria Linn in the southeastern Junggar Basin. Chinese Journal of Plant Ecology , 37 (7) : 665–673. DOI:10.3724/SP.J.1258.2013.00069 [in Chinese] ) ( 0)
|
| [22] |
周雅聃, 陈世苹, 宋维民, 等. 2011. 不同降水条件下两种荒漠植物的水分利用策略. 植物生态学报 , 35 (8) : 789–800.
( Zhou Y D, Chen S P, Song W M, et al.2011. Water-use strategies of two desert plants along a precipitation gradient in northwestern China. Chinese Journal of Plant Ecology , 35 (8) : 789–800. DOI:10.3724/SP.J.1258.2011.00789 [in Chinese] ) ( 0)
|
| [23] |
朱雅娟, 贾志清, 卢琦, 等. 2010. 乌兰布和沙漠5种灌木的水分利用策略. 林业科学 , 46 (4) : 15–21.
( Zhu Y J, Jia Z Q, Lu Q, et al.2010. Water use strategy of five shrubs in Ulanbuh desert. Scientia Silvae Sinicae , 46 (4) : 15–21. [in Chinese] ) ( 0)
|
| [24] |
Ellsworth P Z, Williams D G.2007. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant&Soil , 291 (1/2) : 93–107.
(0)
|
| [25] |
Hasselquist N J, Allen M F, Santiago L S.2010. Water relations of evergreen and drought-deciduous trees along a seasonally dry tropical forest chronosequence. Oecologia , 164 (4) : 881–890.
DOI:10.1007/s00442-010-1725-y (0)
|
| [26] |
Jackson P C, Meinzer F C, Bustamante M.1999. Partitioning of soil water among tree species in a Brazilian Cerrado ecosystem. Tree Physiology , 19 : 717–724.
DOI:10.1093/treephys/19.11.717 (0)
|
| [27] |
Jia G D, Yu X X, Deng W P, et al.2012. Determination of minimum extraction times for water of plants and soils used in isotopic analysis. Journal of Food,Agriculture & Environment , 20 (3/4) : 1035–1040.
(0)
|
| [28] |
Lin G, Phillips S L, Ehleringer J R, et al.1996. Monsoonal precipitation responses of shrubs in a cold desert community on the Colorado Plateau. Oecologia , 106 : 8–17.
DOI:10.1007/BF00334402 (0)
|
| [29] |
Penuelas J, Canadell J G, Ogaya R.2011. Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Global Ecology&Biogeography , 20 (4) : 597–608.
(0)
|
| [30] |
Yang H, Auerswald K, Bai K, et al.2011. Complementarity in water source among dominant species in typical strppe ecosystems of Inner Mongolia,China. Plant and Soil , 340 (1/2) : 303–313.
(0)
|
| [31] |
Zhang W, Cheng B, Hu Z, et al.2010. Using stable isotopes to determine the water sources in alpine ecosystems on the east Qinghai-Tibet plateau,China. Hydrological Processes , 24 (22) : 3270–3280.
DOI:10.1002/hyp.v24:22 (0)
|