 2015, Vol. 51
2015, Vol. 51文章信息
- 刘龙昌, 陶珂, 景康康, 王菲, 司卫杰
- Liu Longchang, Tao Ke, Jing Kangkang, Wang Fei, Si Weijie
- 花色多态植物紫薇花部综合特征与繁育系统
- Floral Syndrome and Breeding Systems of the Flower Color Polymorphic Plant Lagerstroemia indica
- 林业科学, 2015, 51(1): 42-54
- Scientia Silvae Sinicae, 2015, 51(1): 42-54.
- DOI: 10.11707/j.1001-7488.20150105
-
文章历史
- 收稿日期:2014-02-24
- 修回日期:2014-07-15
-
作者相关文章


2. 中国科学院武汉植物园 武汉 430074;
3. 甘肃农业大学草业学院 兰州 730070
2. Wuhan Botanical Garden, Chinese Academy of Sciences Wuhan 430074;
3. College of Pratacultural Science, Gansu Agricultural University Lanzhou 730070
繁育系统是指直接影响后代遗传组成的所有有性特征,主要包括花部综合特征、花部各性器官的寿命、花开放式样、传粉者种类和传粉行为以及自交亲和程度和交配系统(Wyatt,1983; 何亚平等,2003)。花部综合特征与传粉者行为、传粉机制和植物适合度密切相关(李永泉等,2007)。花部形态上的变化会直接影响植物交配系统的各个组成部分,如传粉者的访花行为、自花异花传粉的程度以及雄性或雌性繁殖适合度(吴雪莲等,2007)。对具有花色多态型的植物来说,传粉昆虫的非随机性访花行为会直接影响不同花色植株的交配式样(Romina et al.,2012)。
紫薇(Lagerstroemia indica)是千屈菜科(Lythraceae)紫薇属(Lagerstroemia) 植物,为我国著名夏季观赏花木。在栽培和自然条件下,该植物均具有多种不同花色的植株,其中比较稳定易辨的有白色、红色和紫色花植株(中国科学院中国植物志编辑委员会,1983; 张亚东等,2004; Qin et al.,2007)。从花部形态特征来看,该植物具有6枚与花柱长度和弯曲度类似的长雄蕊和多数簇拥于花中央的短雄蕊,属典型的“异型雄蕊”(任明迅,2009)。丰富的花色及花部形态变化使其成为植物繁育系统多样性研究的良好素材,但目前紫薇在这方面的研究报道还很少,其异型雄蕊在功能上是否发生分化也存在争议。Nepi等(2003)研究表明紫薇2种雄蕊的花粉在颜色、数量、大小、花粉壁结构、生活力以及化学成分上都有一定的差异,已经发生了功能上的分化; 但张秦英等(2008)的研究却显示这2种雄蕊的花粉尚未发生功能上的分化。为此,本文对紫薇的花部特征和繁育特性进行了详细观测,旨在探讨: 1)3种花色植株的花部形态特征是否存在差异; 2)3种花色植株的传粉生物学特性和交配系统是否存在差异; 3)其异型雄蕊在功能和生理上是否存在分化。研究结果也可为紫薇的引种驯化、繁殖和杂交育种工作提供理论依据。
1 材料与方法 1.1 试验材料和研究地点概况紫薇原产亚洲,现广植于热带地区。我国广东、广西、湖南、福建、江西、浙江、江苏、湖北、河南、河北、山东、安徽、陕西、四川、云南、贵州及吉林均有生长或栽培。它适应性强,花朵较大,颜色鲜艳,具有很高的观赏价值。花朵常组成顶生圆锥花序; 雄蕊36~42 枚,外面6枚着生于花萼上,比其余的长得多; 花期6—9月,果期9—12月(Qin et al.,2007)。
野外观察试验于2011年6—10月在洛阳市周山森林公园紫薇栽培种群(34°37′ 37″N,112°22′46″E; 海拔216.93 m)进行。2012年同期进行了验证性观察。研究样地红色、紫色和白色花植株比例为0.6: 0.25: 0.15。各花色植株随机排列,株行距为2 m × 3 m。各花色植株于2001年定植,树龄约为14年。样地地处暖温带季风气候区,气候温和,四季分明。年均气温14.7 ℃,1月均温-2.51 ℃,7月均温25.8 ℃; 年平均日照2 327 h,无霜期210天; 平均降水量601.6 mm,且分布不均,夏季(7,8月份降水最多)降水量占全年降水量的46.9%。
1.2 研究方法 1.2.1 花部形态和开花生物学特性观测在盛花期,随机取3种花色植株各10株,点数每株上的花序数; 每个植株随机选取3个花序,计数每个花序上同时开花的花朵数; 在抽取的花序上随机选取2朵花,观测花径,花瓣长度和宽度,花丝和花药的长度,长、短雄蕊的长度,花柱长度,子房长度以及短雄蕊数量和雄蕊群面积。其中花药和子房的长度在解剖镜下测量,其余指标用游标卡尺测量。雄蕊群面积=雄蕊群长度×雄蕊群宽度(紫薇花辐射对称,长宽大致相等)。
随机选取3种花色植株各10~15株,对即将开放的花序进行标记,观察其单个花序的开花特性。每株选取3朵第2天即将开放的单花进行标记,观察其单花的开花特性,从花开前1天10:00开始,每隔0.5 h观察记录所标记花的开放状态。花开放后每天上午9:00观察1次直至花瓣脱落为止。
1.2.2 花粉组织化学检测参照Dafni(1992)的方法。随机选取3种花色植株各10株,每株上取1个花序,每个花序上取1朵刚开放的花,分别取其长、短雄蕊的成熟花粉置于凹面载玻片上,加1滴I2-KI或苏丹III-IV染色液,在显微镜下观察花粉粒的染色情况。
1.2.3 单花花粉量、胚珠数和花粉/胚珠比率(P/O)测定参照路国辉等(2009)的方法进行。随机选取3种花色植株各10株,每株取3朵刚开放而花药尚未开裂的花,固定于FAA中,带回实验室测定。
1.2.4 花粉败育率测定参照陈家瑞(l991)的孢粉染色法。随机选取3种花色植株各10株,每株选3个花序,每花序选1朵刚开放的花,分别取其长、短雄蕊的花粉置于不同载玻片上,加1滴孔雀绿-酸性品红-橘红G染液,盖上盖玻片,在显微镜下观察统计可育和败育的花粉数。
1.2.5 花粉活力的测定采用TTC法(Dafni,1992)。随机选取3种花色植株各10株,每株选1 个花序,每花序选1朵花,自长、短雄蕊花药开裂时起每隔2 h分别取其花粉置于不同载玻片上,用TTC染液进行染色,在40 ℃的培养箱中染色15~30 min后,置光学显微镜下观察。
1.2.6 柱头可授性测定参照吴雪莲等(2007)的方法。随机选取3种花色植株各10株,随机标记发育阶段一致的花,从花开放第1天7:00开始每隔2 h取10朵花的柱头(每株取1朵),浸入含有联苯胺-过氧化氢反应液(联苯胺: 过氧化氢: 水=4: 11: 22)的凹面载玻片中,盖上盖玻片,置于显微镜下观察,至19:00。此后每天9:00测定1 次,直至所标记的花凋落。
1.2.7 访花昆虫及其行为观察随机选择3种花色植株各3株,每株上选取10朵新开的花进行挂牌,连续观测3天。从7:00—17:00,连续观察记录访花昆虫种类、访花频率和访花行为,并捕捉昆虫制作标本。访问频率为每朵花在1 h内被访问的次数,不同时段访花频率的平均数为平均访花频率。
在靠近样地中心的位置设置10 m×10 m样方,样方中红色花、紫色花和白色花植株分别为6,3,3株。随机选择3种花色植株各2株,从7:00—17:00,连续观察记录传粉昆虫的访问次数,连续观察3天,取平均值为某一花色植株访问次数。
访问次数理论值即期望值,假设访花昆虫对不同花色植株的访问是随机的、花展示面积依赖的,则某花色植株访问次数期望值=实际访问次数(3种花色之和)× 该花色植株所占比例 × 该花色植株花序总面积系数。单花序面积=花序长 × 花序宽 ÷ 2; 植株花序总面积=单花序面积 × 花序数。
1.2.8 不同花部构件对传粉昆虫招引作用的检测随机选取3种花色植株各10~15株,每株分如下处理:1)自然对照; 2)去除暗黄色长雄蕊; 3)去除鲜黄色短雄蕊;4)去除全部雄蕊; 5)去除花瓣。然后挂牌,直至结果。统计各项处理的结实率和结籽数,同时,检测上述几种处理条件下传粉昆虫的访花频率(方法同1.2.7)。以上所有处理是在单花上进行的。结实率=果实数/花数×100%。
1.2.9 人工控制授粉试验随机选取3种花色植株各10~15株,每株分如下处理:1)自然对照; 2)花开前套袋,直至花凋谢; 3)花开前套袋,花开后人工自花授粉; 4)花开前套袋,花开后人工异花授粉。检测各处理的结实率。
1.3 数据分析对花粉败育率等百分比数据进行反正弦转换,对单花花粉量等差异较大的数据进行自然对数转换,以满足方差分析的要求。用单因素方差(One Way ANOVA)法对3种花色植株的花部特征、花粉量和花粉/胚珠(P/O)、花粉活力和败育率、访花频率以及结实率和结籽数等进行方差分析,用Tukey HSE法进行显著性检验; 用拟合优度检验(Chi-square goodness-of-fit test)法对传粉昆虫的访花偏好进行分析(Romina et al.,2012)。用SPSS 18.0软件包对所测数据进行处理分析,用Excel软件作图。
2 结果与分析 2.1 紫薇花部形态特征及其开花特性紫薇红色花植株的花序数以及同时开放的花朵数最多,花径最大,花药(长、短雄蕊花药)也最长,与紫色花或白色花植株存在显著差异。3种花色植株的花序面积差异不显著。3 种花色植株的花柱长和短雄蕊花丝长差异均不显著。红色花植株的雄蕊群面积最小,长雄蕊花丝最短,与紫色花和白色花植株存在显著差异。红色花植株的花瓣最宽,明显宽于紫色花和白色花植株的。而紫色花植株的花瓣最长,明显长于红色和白色花植株的。3种花色植株的长雄蕊数均为6枚,所以,表 1中雄蕊数的差异实际上是短雄蕊数的差异。白色花植株的短雄蕊数最多,红色花植株的短雄蕊数最少(表 1)。3种花色植株的长雄蕊花丝和花药长度均显著高于短雄蕊的(P<0.01)。
|
|
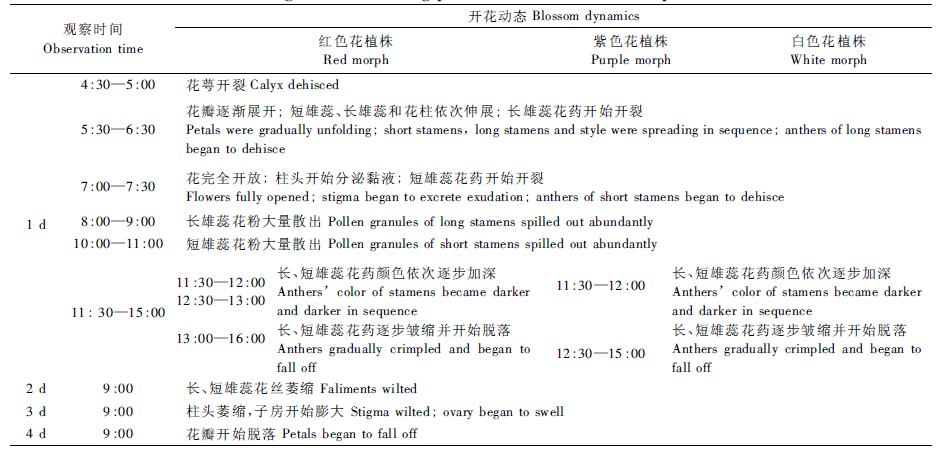
在研究地点,紫薇3种花色植株的花期均可从6月底持续到10月初。3种花色植株单个圆锥花序的开花持续时间大体一致,均为50天左右。3种花色植株的单花开花进程也相似,单花花期为4~5天。晴天花期较短,为4天; 阴天花期较长,为5天。在晴天,花萼于清晨4:30左右开裂,花7:00左右完全开放。它们的花药开始和完全开裂(大量散粉)的时间相同,但红色花植株的花药形态和颜色维持时间略长于紫色和白色花(约1 h)。它们的长雄蕊均比短雄蕊早1 h开裂(表 2)。花开放后约3 h,长雄蕊花药外围出现褐圈,比短雄蕊花药变色稍早(1 h左右)。花开放后5~6 h 2种花药开始逐步变干皱缩。48 h后子房膨大、柱头蜷缩。依天气的不同,花瓣于其后2~3天逐渐枯萎脱落(表 2)。
|
|
白色和紫色花植株的单花花粉量和P/O值相近,均显著高于红色花植株(P<0.01)。但它们的胚珠数差别不大(表 3)。
|
|
3种花色植株长、短雄蕊之间的花粉量均存在极显著差异(P<0.01),短雄蕊的花粉量分别是长雄蕊的约3.2倍、3.0倍和2.55倍。但单个短雄蕊的花粉量要远低于长雄蕊。
2.3 花粉的组织化学成分和花粉败育率3种花色植株长、短雄蕊之间的花粉组织化学成分没有差异,都是既含淀粉又有脂质(表 4)。红色花植株的长、短雄蕊花粉败育率最高,显著高于其他2种花色(P<0.05)。各花色植株长、短雄蕊之间的花粉败育率接近,差异均不显著(表 5)。
|
|
|
|
3种花色植株的花粉活力变化趋势一致(表 6),花药刚开裂时,长、短雄蕊的花粉已有较强活力,均达40%以上。长、短雄蕊花粉活力均在大量散粉时达到最高,此时它们的花粉活力无明显差异。长雄蕊花粉活力在花开后6 h内可维持较高水平,而短雄蕊的能维持8 h。
|
|
3种花色植株的柱头可授性变化趋势比较一致(表 6),花开放时柱头已具有可授性,花开后4 h可授性达到最佳。红色花植株的柱头最佳可授期维持时间最短(约6 h); 紫色和白色花植株的最佳可授期均能维持约8 h; 紫色花植株柱头可授期最长(约50 h)。3种花色植株的花粉寿命与柱头可授期均有重叠,短雄蕊花粉活力最高时期与柱头最佳可授期比较同步,长雄蕊花粉活力最高时期早于柱头最佳可授期。
2.5 传粉昆虫及其行为紫薇3 种花色植株的访花昆虫基本一致,主要有黄胸木蜂(Xylocopa appendiculata)、中华蜜蜂(Apis cerana)、意大利蜜蜂(A. mellifera)、切叶蜂(Megachile sp.)、食蚜蝇(Syrphidae sp.)、芦蜂(Ceratina sp.)和隧蜂(Halictus sp.),偶见长脚蜂(Polistes sp.)、蝇类及甲虫等无规律访花(图 1)。
 |
图 1 紫薇的传粉昆虫 Fig. 1 Pollinators of L. indicaA, B. 黄胸木蜂Xylocopa appendiculata; C.切叶蜂Megachile sp.; D.芦蜂Ceratina sp.; E. 食蚜蝇Syrphidaesp.; F. 意大利蜜蜂 Apis mellifera. |
黄胸木蜂是最主要的传粉昆虫,其访花行为很有规律,总是从花的正面直奔黄色的短雄蕊,降落到短雄蕊上取食短雄蕊花粉。在起飞、降落和取食过程中,高频振动的身体使长、短雄蕊的花粉弹到其背腹部及身体两侧。在此过程中,其身体也会与外轮长雄蕊的花药或柱头接触。这样在访问下一朵花时,花粉就被带到下一朵花的柱头上。蜜蜂、切叶蜂和芦蜂的访花行为与黄胸木蜂相同。食蚜蝇既取食短雄蕊花粉,也取食长雄蕊花粉(此时,将身体悬停于空中)。但由于它们的体型较小,与长雄蕊和柱头直接接触的机会很少,身体上携带的花粉数量也有限,所以其传粉的有效性可能会低于黄胸木蜂; 隧蜂的访花行为也与黄胸木蜂相同,但由于其体型很小,所以可能不是有效的传粉者。
传粉昆虫的平均访花频率为(1.59±0.38)times·flower-1h-1(3种花色植物平均值),最有效的传粉者为黄胸木蜂,其访花次数占总访花次数的51.96%。黄胸木蜂在访花时常连续访问同花色的多个植株或同一植株上的多朵花(可达20朵),可在同一植株的花序间往返多次,有时一朵花可被连续访问2次以上。黄胸木蜂访花高峰出现在8:00—10:30,访花次数可达(1.18±0.37)times·flower-1h-1,此后访花频率逐渐降低,12:00以后,访花次数降至(0.1±0.04)times·flower-1h-1; 其他传粉昆虫到15:00还很活跃。18:00以后不再有传粉昆虫访花。
黄胸木蜂体型大,受天气影响较小,阴天仍可保持一定的访花频率(0.68±0.11 times·flower-1h-1)。其他传粉昆虫受天气影响较大,在阴雨天或大风天,访花频率很低或不访花。
3种花色植株的有效传粉者的访花高峰期基本一致,均为双峰型: 09:00—10:00和15:00—16:00。拟合优度检验表明,传粉昆虫对红色花植株有明显偏好(表 7)。传粉昆虫访问红色花植株的频率显著高于紫色和白色花(P<0.01),紫色和白色花植株间无明显差异(图 2,表 8)。
|
|
 |
图 2 传粉昆虫日访花频率 Fig. 2 Daily visiting frequency of pollinators of the three floral morphs of L. indica |
|
|
3种花色植株在5种处理下的访花频率、结实率和结籽数呈现一致的变化。去除短雄蕊后,访花频率、结实率和结籽数均显著低于对照(P<0.01),说明黄色短雄蕊起招引传粉昆虫的作用; 去除长雄蕊后,访花频率显著高于对照,结实率和结籽数虽明显高于去除短雄蕊的处理,但仍显著低于对照,说明暗黄色长雄蕊没有招引传粉昆虫的作用,同时,也暗示长雄蕊起传粉作用,可能为传粉型雄蕊; 去全部雄蕊后,不再有传粉昆虫访花,也不再结实,进一步说明黄色短雄蕊对传粉昆虫有招引作用; 去除花瓣后,访花频率、结实率和结籽数与对照没有显著差异,但因本试验是在单花水平上进行的,所以还不能明确花瓣对传粉昆虫是否有招引作用。
2.7 人工控制授粉试验3种花色植株花前套袋处理均未见结实,说明它们没有主动自交机制,也不存在无融合生殖。用短雄蕊花粉或长雄蕊花粉自交和异交,其结实率间没有明显差异(表 9),表明这2种雄蕊花粉的雄性功能没有显著差异。紫色和白色花植株人工授粉后的结实率显著高于自然结实率(表 9)。
|
|
紫薇单株花序数可多达50个以上,单个花序同时开花数25朵以上(红色、紫色、白色花植株分别为40.62,30.15,28.75),单个花序花期约50天,单花花期为4~5天,说明紫薇具有花展示大、花期长、同步大量开花的开放特征。在花期,吸引了很多传粉昆虫频繁访花,这有利于其花粉的顺利输出和异交的发生;同时,其主要传粉昆虫在访花时常连续访问同一植株上的多朵花,这又不可避免地导致同株异花传粉和自交的发生。此外,紫薇3种花色植株的P/O值分别为1 753.80±172.90,2 021.36±240.60和2 034.19±175.98,按照Cruden(1977)的标准,其交配方式应属于以异交为主、自交亲和的兼性异交繁育系统。套袋试验也很好地证实了这一点,同时,套袋试验还表明紫薇不存在无融合生殖和主动自交现象。综上所述,紫薇的繁育系统是被动自交和异交混合交配系统。因近交衰退使近交的便利减小,而远交的代价很高,受外界条件影响也大,所以,权衡近交和远交的利弊,折中的方案是混合的交配系统(何亚平等,2003)。这种交配系统既能产生遗传多样性丰富的远交后代也能产生遗传性状稳定不变的近交后代(刘芬等,2013)。
尽管如此,紫薇3种花色植株的交配系统可能存在一定差异。与其他2种花色植株相比,红色花植株的花序数和同时开放的花朵数更多一些,也吸引了更多的传粉昆虫(访问频率高),所以其同株异花授粉引起的自交率可能也会更高一些(Harder et al.,1995)。
3.2 紫薇花部特征与传粉的适应性多花的开花式样和大花径比少花的开花式样和小花径更为醒目,能吸引更多的传粉者访花(Podolsky,1992; Brunet et al.,2006),从而导致较高的座果和花粉输出率(Ishii et al.,2002; 王茜等,2012)。而花蜜和花粉可能在近距离对传粉者有诱导作用(黄双全等,2000)。紫薇没有蜜腺,其鲜艳的花瓣特别是簇拥在花中央的鲜黄色短雄蕊,吸引了传粉昆虫为其传粉。色彩鲜艳的花药在访花者眼里就像一个醒目的广告,对于那些以花粉为食物的传粉者尤其具有诱惑力(任明迅,2009)。本研究表明,与其他2种花色植株相比,红色花植株的花序数和同时开放的花数最多,花径最大,花药最长,黄色短雄蕊与花瓣的颜色对比也最为鲜明。因此,在自然条件下,传粉昆虫访问该花色植株的频率显著高于其他2种花色植株(P<0.01)。此外,红色花植株的长雄蕊花丝最短(表 1),与短雄蕊的间距最小,这增加了传粉昆虫(尤其体型较大的黄胸木蜂)背部与位于上部的长雄蕊花药和柱头接触的机会。同时,传粉昆虫在起降和取食短雄蕊花粉时,为了躲避外围长雄蕊的干扰,会更靠近位于中部的雌蕊,这又增加了给柱头传粉的机会。因此,红色花植株的结实率和结籽数会显著高于其他2种花色植株。然而,紫色花和白色花植株的短雄蕊数和雄蕊群面积却显著大于红色花植株,因而有利于吸引近距离的传粉者访花,这在一定程度上弥补了其花展示的不足,保障了繁殖的成效。另外,紫薇的花柱先端呈钩状向上弯曲,这增大了柱头接受传粉昆虫体表花粉的机会。对花径较小的紫色花和白色花植株来说,因传粉昆虫活动的空间有限,其柱头接受传粉昆虫体表花粉的机会可能也会更高一些。
本研究表明,紫色和白色花植株人工授粉后的结实率显著高于自然对照,表明这2种花色植株存在一定的花粉限制,传粉媒介不足很可能是影响其结实率的主要原因(刘芬等,2013)。与红色花植株相比,它们的单花雄蕊数、花粉数和P/O值明显高出许多,花粉活力也高一些,表现出偏雄的性分配(Ishii,2002; 马晓丽等,2011)。这有助于减轻由于传粉者不足造成的不利因素的影响,可能是对花粉限制的有效补偿(Guo et al.,2010)。偏雄的性分配有助于其在花期通过输出花粉提高雄性适合度,从而实现个体适合度的最大化(戈星月等,2012)。
大量研究表明,许多植物的花在实现雌性和雄性繁殖功能后,寿命会显著缩短或很快终止(Van Doorn,1997; Evanhoe et al.,2002; Castro et al.,2008)。但紫薇的花瓣在柱头和花药萎蔫后仍可维持1~2天的新鲜状态,这既增加了其植株水平上的花展示,有利于雄性适合度的提高,同时也不会因花展示的增加而同步扩大同株异花授粉的机会。因此,紫薇通过花瓣寿命延长增加花展示是有利的,是一种积极的适应策略(彭德力等,2012)。
花最主要的诱物包括颜色、大小、形状和气味,其中花色是植物吸引传粉者的最重要视觉信号(Menzel et al.,1993; 张大勇,2004)。许多研究显示,传粉者访花偏好性与花色有关(施海燕等,2008; Romina et al.,2012)。颜色鲜艳的花对传粉者的吸引力更强,往往比白色花具有更高的异交率和适合度(吴雪莲等,2007; Potts et al.,2010)。本研究表明,传粉昆虫访问红色花植株的频率显著高于其他2种花色植株,对红色花有明显偏好(表 7)。这应该是红色花植株自然授粉结实率和结籽率显著高于后二者的主要原因。既然如此,种内花色多态性又是如何维持的呢?连续访问同色花(colour constancy,花色忠实性)虽然可以减少传粉昆虫在不同花色间选择和更替(随机性)带来的成本,但也增加了因重复访问相同的花而带来的收益减少的风险,在花色忠实性和随机性之间存在权衡关系,当花提供的报酬较多且花间隔距离较远时,传粉昆虫倾向于随机性访花(即访花忠实性降低)(Gegear et al.,2004)。随机性访花可能是花色多态性得以维持的原因(Romina et al.,2012)。颜色鲜艳的花能吸引更多的传粉者,同样也会引来更多植食者,因而可能是传粉者和植食者共同维持了花色的多态性(Strauss et al.,2004)。另外,花色多态性还可能与土壤水分有直接关系。含有色素的植株在干旱的环境下表现更好,而无色素的植株则在水分充足的条件下具有更高的适合度(Warren et al.,2001; Tang et al.,2010)。上述研究表明,花色多态性的维持和演化可能受到多种因子共同影响。
在本研究中,花瓣对传粉昆虫招引作用的观测是在单花水平上进行的。对于一个有25个以上花朵同时开放的圆锥花序来说,仅去除1朵花的花瓣对传粉昆虫吸引力的影响十分有限。另外,在近处,对传粉昆虫起吸引作用的往往是花粉(彭东辉等,2008)。也就是说,传粉昆虫在决定访问花序中的哪朵花时,可能主要受花粉的影响。这可能是去除单花花瓣对传粉昆虫访问无显著影响的主要原因。从传粉昆虫对红色花有明显偏好这一研究结果来看,花瓣对传粉昆虫应该有一定招引作用。为进一步探明紫薇花瓣对传粉昆虫招引作用,在单花、花序以及单株等多个水平上同时开展对比研究是十分必要的。
3.3 紫薇异型雄蕊在传粉过程中的功能分化植物以雄蕊的花药和花粉来吸引传粉者或作为传粉者的回报物,或以花药颜色作为吸引传粉者的指示物(Ashman,2000; Lau et al.,2004),其形态与颜色的变化会影响花的吸引力和访花者的访花行为(Harder et al.,1993; Kudo,2003)。
紫薇2种雄蕊的花药颜色以及花药、花丝长度差异明显,2种雄蕊的花粉在数量、大小以及花粉壁结构上也有一定差异(Nepi et al.,2003),这是2种雄蕊功能分化的形态结构基础。本研究表明,绝大多数传粉昆虫在访花时,会直接飞向花中心的黄色短雄蕊,而不理会外围的暗黄色的长雄蕊。去除黄色短雄蕊后,访花频率、结实率和结籽数均显著低于自然对照; 去除长雄蕊后,访花频率甚至显著高于对照。据此推断紫薇的2种雄蕊在传粉过程中存在一定程度的功能分化,黄色短雄蕊主要起招引昆虫作用,暗黄色的长雄蕊主要起传粉作用。研究结果还显示,食蚜蝇对2种雄蕊并未表现出明显的选择性,2种雄蕊的花粉都会采集,因此,异型雄蕊的功能分化也许还与传粉昆虫的种类有关(路国辉等,2009)。去除长雄蕊后,传粉昆虫的访花频率虽显著高于对照,但结实率和结籽数却未同步提高。这可能与其独特的雄蕊排列方式有关,即向内弯曲的长雄蕊位于花的外轮,短雄蕊位于花的中央。去除外轮长雄蕊为传粉昆虫进入花心取食短雄蕊清除了障碍,所以访花频率会提高;同时,去除长雄蕊后,传粉昆虫可以更方便地从侧面进出花心,从而大大减少了传粉昆虫与柱头接触或柱头接受花粉的机会,这可能是其结实率和结籽数未能同步提高的主要原因。同理可知,紫薇长、短雄蕊的这种排列方式可能也是二者在功能上出现一定分化的原因之一。
在大多数具有异型雄蕊的植物中,吸引传粉者的雄蕊(给食型)往往花粉活力很低、花粉数量较少甚至没有花粉,而传粉型雄蕊往往花粉活力较高、花粉数量较多(Bowers,1975; Escaravage et al.,2001; Luo et al.,2008)。这是植物在资源有限条件下提高雄蕊对传粉者吸引能力的有效方式,可以使植物以最小的代价获得较高的适合度(Jesson et al.,2003; 路国辉等,2009)。本研究表明,紫薇2种雄蕊的花粉活力在散粉高峰期没有明显差异,二者的组织化学成分以及人工控制授粉后的结实率也没有显著的差异。2类花粉在花开放的6 h内都有很高的活力,均可作为传粉型雄蕊;且短雄蕊的总花粉量也显著高于长雄蕊的。这表明2种雄蕊的花粉在生理功能上尚未分化。与Nepi等(2003)的研究结果不同,本文的研究进一步证实了张秦英等(2008)的研究结论。另外,本研究还表明,紫薇2种雄蕊花药的开裂时间不同步(短雄蕊的花药开裂滞后于长雄蕊花药1~2 h),短雄蕊花粉活性维持的时间长于长雄蕊花粉。花内雄蕊的这种分化使得不同雄蕊的花粉逐步成熟并分批呈给传粉者,提高了花粉的散布效率,这是植物适应蜂类传粉和最大化输出花粉的一种适应机制(廖万金等,2007; 陆婷等,2007)。
尽管有许多研究支持“吸引传粉者雄蕊的花粉数量较少甚至没有花粉,而传粉型雄蕊的花粉数量较多”这一结论。但也有不少不同的研究报道,例如,Dulberger(1981)的研究表明,耳叶决明(Cassia auriculata)每个长雄蕊(传粉型)和短雄蕊(给食型)的花粉数量大体相等,4枚短雄蕊的总花粉量明显高于3枚长雄蕊的;又如,刺萼龙葵(Solanum rostratum)单个给食型雄蕊的花粉量虽然低于单个传粉型雄蕊的花粉量,但是给食型雄蕊(4枚)的总花粉量却显著高于传粉型雄蕊(1枚)的总花粉量(Vallejo-Marín et al.,2009)。这与本文的研究结果一致。Gross等(2001)认为,传粉昆虫对给食型雄蕊的偏好可能是由于给食型雄蕊的资源(花粉)相对更为丰富一些。以上研究表明,异型雄蕊的分工可能并不像人们想象的那么简单,它可能是一个非常复杂的问题,不同植物分类群的异型雄蕊在分工程度和方式上可能有很大的差异,弄清这些问题还需要更为系统和深入的研究(罗中莱等,2005; 路国辉等,2009)。
| [1] |
陈家瑞.1991. 植物孢粉染色技术综述及其应用. 植物学集刊, (5):269-276. (Chen J R. 1991. Review on the staining methods of plant spore and its application. Botanical Research, (5): 269-276.[in Chinese])(  0) 0)
|
| [2] |
戈星月,朱璧如,廖万金. 2012. 雌全同株植物三脉紫菀花期偏雄的个体大小依赖的性别分配. 生物多样性, 20 (3):386-390. (Ge X Y, Zhu B R, Liao W J. 2012. Male biased sex allocation with plant size in gynomonoecious Aster ageratoides. Biodiversity Science, 20 (3):386-390[in Chinese]).( 1)
|
| [3] |
何亚平,刘建全. 2003. 植物繁育系统研究的最新进展和评述. 植物生态学报, 27(2):151-163. (He Y P, Liu J Q. 2003. A review on recent advances in the studies of plant breeding system. Acta Phytoecologica Sinica, 27(2):151-163[in Chinese]).( 2)
|
| [4] |
黄双全,郭友好. 2000.传粉生物学的研究进展. 科学通报, 45(3):225-237. (Huang S Q, Guo Y H. 2000. New advances in pollination biology and studies in China. Chinese Science Bulletin, 45(3):225-237[in Chinese]).( 1)
|
| [5] |
李永泉,张奠湘. 2007. 山地五月茶的蝇类传粉研究. 植物分类学报, 45 (2):217-226. (Li Y Q, Zhang D X. 2007. Fly pollination of Antidesma montanum (Euphorbiaceae) in Hainan, China. Acta Phytotaxonomica Sinica, 45 (2):217-226[in Chinese]).( 1)
|
| [6] |
廖万金,王峥媚,谢丽娜,等. 2007. 草乌传粉过程中的广告效应与回报物质研究. 生物多样性, 15(6): 618-625. (Liao W J, Wang Z M, Xie L N, et al. 2007. Floral advertisement and rewards in bumblebee-pollinated Aconitum kusnezoffii (Ranunculaceae). Biodiversity Science, 15(6): 618-625[in Chinese]).( 1)
|
| [7] |
刘芬,李全健,王彩霞,等. 2013. 濒危植物扇脉杓兰的花部特征与繁育系统. 林业科学, 49(1):53-60. (Liu F, Li Q J, Wang C X, et al. 2013. Floral characteristics and breeding systems of an endangered species Cypripedium japonicum. Scientia Silvae Sinicae, 49(1):53-60[in Chinese]).( 2)
|
| [8] |
陆婷,谭敦炎. 2007. 动物传粉植物花粉呈现时序的进化意义. 生物多样性, 15(6):673-679. (Lu T, Tan D Y. 2007. Evolutionary implications of pollen presentation schedules in animal-pollinated plants. Biodiversity Science, 15(6):673-679[in Chinese]).( 1)
|
| [9] |
路国辉,武文华,王瑞珍,等. 2009. 野牡丹异型雄蕊的功能分化. 生物多样性, 17 (2):174-181. (Lu G H, Wu W H, Wang R Z, et al. 2009. Division of labor of heteromorphic stamens in Melastoma malabathricum. Biodiversity Science, 17 (2):174-181[in Chinese]).( 4)
|
| [10] |
罗中莱,张奠湘. 2005. 异型雄蕊的研究进展. 热带亚热带植物学报,13(6):536-542. (Luo Z L, Zhang D X. 2005. A review of heteranthery in flowering plants. Journal of Tropical and Subtropical Botany, 13(6):536-542[in Chinese]).( 1)
|
| [11] |
马晓丽,谭敦炎,李新蓉. 2011. 蒙古沙冬青花序内性分配的变化、传粉者运动与繁殖成功. 生物多样性, 19(4):432-440. (Ma X L, Tan D Y, Li X R. 2011. Variation in floral sex allocation, pollinator movement and reproducetive success in Ammopiptanthus mongolicus inflorescences. Biodiversity Science, 19 (4):432-440[in Chinese]).( 1)
|
| [12] |
彭德力,张志强,牛洋,等. 2012.高山植物繁殖策略的研究进展. 生物多样性, 20 (3): 286-299. (Peng D L, Zhang Z Q, Niu Y, et al. 2012. Advances in the studies of reproductive strategies of alpine plants. Biodiversity Science, 20 (3): 286-299[in Chinese]).( 1)
|
| [13] |
彭东辉,张启翔,黄启堂,等. 2008. 多花野牡丹传粉生物学观察. 福建林学院学报, 28(2):115-120. (Peng D H, Zhang Q X, Huang Q T, et al. 2008.Study on pollination biology of Melastoma affine. Journal of Fujian College of Forestry, 28(2):115-120[in Chinese]).( 1)
|
| [14] |
任明迅. 2009. 花内雄蕊分化及其适应意义. 植物生态学报, 33(1): 222-236. (Ren M X. 2009. Intrafloral stamen differentiations and their adaptive significances. Chinese Journal of Plant Ecology, 33(1): 222-236[in Chinese]).( 2)
|
| [15] |
施海燕, 吴杰, 李继莲, 等. 2008. 小峰熊蜂访花偏爱性. 昆虫学报, 51(9): 946-952. (Shi H Y, Wu J, Li J L, et al. 2008. Foraging preference of the bumblebee Bombus hypocrita (Hymenoptera:Apidae). Acta Entomologica Sinica, 51(9):946-952[in Chinese]).( 1)
|
| [16] |
王茜,邓洪平,丁博, 等. 2012. 钝叶柃不同性别花的花部形态与传粉特征比较. 生态学报, 32(12):3921-3930. (Wang Q, Deng H P, Ding B, et al. 2012. Comparison of floral morphology and pollination characteristics between the sexes in Eurya obtusifolia. Acta Ecologica Sinica, 32(12):3921-3930[in Chinese]).( 1)
|
| [17] |
吴雪莲,谭敦炎. 2007. 异果芥的花部综合征及其繁育系统. 植物分类学报, 45(4):538-550. (Wu X L, Tan D Y. 2007. Floral characters and breeding systems in the dimorphic annual plant Diptychocarpus strictus (Cruciferae). Acta Phytotaxonomica Sinica, 45(4):538-550[in Chinese]).( 3)
|
| [18] |
张大勇. 2004. 植物繁殖生态学//张大勇. 植物生活史进化与繁殖生态学. 北京: 科学出版社, 148-164. (Zhang D Y. 2004. Plant reproductive ecology//Zhang D Y. The evolution of life-history and reproductive ecology of plants. Beijing: Science Press, 148-164.[in Chinese])( 1)
|
| [19] |
张秦英,罗凤霞,刘莉,等. 2008. 紫薇异型雄蕊花粉生活力研究. 园艺学报, 35(12):1741. (Zhang Q Y, Luo F X, Liu L, et al. 2008. Pollen viability of dimorphic anthers in Lagerstroemia indica. Acta Horticulturae Sinica, 35(12):1741[in Chinese]).( 2)
|
| [20] |
张亚东,杨彦伶. 2004. 湖北保康不同花色野生紫薇的RAPD分析. 分子植物育种, 2 (5): 683-688. (Zhang Y D, Yang Y L. 2004. RAPD analysis in the different color-flowered Lagerstroemia indica from Baokang County in Hubei Province. Molecular Plant Breeding, 2 (5): 683-688[in Chinese]).( 1)
|
| [21] |
中国科学院中国植物志编辑委员会.1983. 中国植物志: 第52卷第2分册. 北京: 科学出版社, 92 -111. (China Botanical Biography Compiling Committee, CAS. 1983. China botanical biography: Vol. 52, Sect 2. Beijing: Science Press, 92 -111.[in Chinese])( 1)
|
| [22] |
Ashman T L.2000. Pollinator selectivity and its implication for the evolution of dioecy and sexual dimorphism.Ecology, 81(9):2577-2591.(1)
|
| [23] |
Bowers K A W. 1975. The pollination ecology of Solanum rostratum (Solanaceae). American Journal of Botany, 62(6): 633-638.(1)
|
| [24] |
Brunet J, Sweet H R.2006. Impact of insect pollinator group and floral display size on outcrossing rate. Evolution, 60(2):234-246.(1)
|
| [25] |
Castro S, Silveira P, Navarro L. 2008. Effect of pollination on floral longevity and costs of delaying fertilization in the out-crossing Polygala vayredae Costa (Polygalaceae). Annals of Botany, 102(6):1043-1048.(1)
|
| [26] |
Cruden R W. 1977. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution, 31(1):32-46.(1)
|
| [27] |
Dafni A.1992. Pollination ecology: A practical approach. London: Oxford University Press, 59-89.(2)
|
| [28] |
Dulberger R. 1981. The floral biology of Cassia didymobotrya and C. auriculata (Caesalpinoideae). American Journal of Botany, 68(10):1350-1360.(1)
|
| [29] |
Escaravage N, Flubacker E, Pornon A, et al. 2001. Stamen dimorphism in Rhododendron ferrugineum (Ericaceae): development and function. American Journal of Botany, 88(1): 68-75.(1)
|
| [30] |
Evanhoe L, Galloway L F. 2002. Floral longevity in Campanula americana (Campanulaceae): a comparison of morphological and functional gender phases. American Journal of Botany, 89(4): 587-591.(1)
|
| [31] |
Gegear R J, Thomson J D. 2004. Does the flower constancy of bumble bees reflect foraging economics? Ethology, 110(10):793-805.(1)
|
| [32] |
Gross C L, Kukuk P F. 2001. Foraging strategies of Amegilla anomola at the flowers of Melastoma affine—no evidence for separate feeding and pollinating anthers. Acta Horticulturae, 561: 171-178.(1)
|
| [33] |
Guo H, Mazer S J, Du G Z. 2010. Geographic variation in primary sex allocation per flower within and among 12 species of Pedicularis (Orobanchaceae): proportional male investment increases with elevation. American Journal of Botany, 97(8):1334-1341.(1)
|
| [34] |
Harder L D, Barrett S C H.1993. Pollen removal from tristylous Pontederia cordata: effects of anther position and pollinator specialization. Ecology, 74(4):1059-1072.(1)
|
| [35] |
Harder L D, Barrett S C H.1995. Mating cost of large floral displays in hermaphrodite plants. Nature, 373:512-515.(1)
|
| [36] |
Ishii H S, Sakai S. 2002. Temporal variation in floral display size and individual floral sex allocation in racemes of Narthecium asiaticum (Liliaceae). American Journal of Botany, 89(3):441-446.(2)
|
| [37] |
Jesson L K, Kang J, Wagner S L, et al. 2003. The developmental basis of enantiostyly. American Journal of Botany, 90(2): 183-195.(1)
|
| [38] |
Kudo G. 2003. Anther arrangement influences pollen deposition and removal in hermaphrodite flowers. Functional Ecology, 17(3):349-355.(1)
|
| [39] |
Lau J A, Galloway L F. 2004. Effects of low-efficiency pollinators on plant fitness and floral trait evolution in Campanula americana (Campanulaceae). Oecologia, 141(4):577-583.(1)
|
| [40] |
Luo Z, Zhang D, Renner S S. 2008. Why two kinds of stamens in buzz-pollinated flowers? Experimental support for Darwin's division-of-labour hypothesis. Functional Ecology, 22(5):794-800.(1)
|
| [41] |
Menzel R, Shmida A. 1993. The ecology of flower colours and the natural colour vision of insect pollinators: the Israeli flora as a study case. Biological Review, 68(1): 81-120.(1)
|
| [42] |
Nepi M, Guarnieri M, Pacini E. 2003. "Real" and feed pollen of Logerstroemia indica ecophysiological diferences. Plant Biology, 5(3): 3l1-314.(3)
|
| [43] |
Podolsky R D. 1992. Strange floral attractors: pollinators attraction and the evolution of plant sexual systems. Science, 258:791-793.(1)
|
| [44] |
Potts S G, Biesmeijer J C, Kremen C, et al. 2010. Global pollinator declines: trends, impacts and drivers. Trends in Ecology & Evolution, 25(6):345-353.(1)
|
| [45] |
Qin H N, Shirley G. 2007. Lythraceae // Wu Z Y, Raven P H, Hong D Y. Flora of China: Vo1.13. Beijing:Science Press, 277-281.(2)
|
| [46] |
Romina M, Julieta N. 2012. Pollinator response to flower color polymorphism and floral display in a plant with a single-locus floral color polymorphism: consequences for plant reproduction. Ecological Research, 27(2): 377-385.(4)
|
| [47] |
Strauss S Y, Irwin R E, Lambrix V M. 2004. Optimal defence theory and flower petal colour predict variation in the secondary chemistry of wild radish. Journal of Ecology, 92(1):132-141.(1)
|
| [48] |
Tang X X, Huang S Q. 2010. Fluctuating selection by water level on gynoecium colour polymorphism in an aquatic plant. Annals of Botany, 106(5): 843-848.(1)
|
| [49] |
Vallejo-Marín M, Manson J S, Thomson J D, et al. 2009. Division of labour within flowers: heteranthery, a floral strategy to reconcile contrasting pollen fates. Journal of Evolutionary Biology, 22(4): 828-839.(1)
|
| [50] |
Van Doorn W G.1997. Effects of pollination on floral attraction and longevity. Journal of Experimental Botany, 48(9):1615-1622.(1)
|
| [51] |
Warren J, Mackenzie S. 2001. Why are all colour combinations not equally represented as flower-color polymerphisms? New Phytologist, 151(1): 237-241.(1)
|
| [52] |
Wyatt R. 1983. Pollinator-plant interactions and the evolution of breeding systems // Real L. Pollination biology. Orlando: Academy Press, 51-95(1)
|