 2014, Vol. 50
2014, Vol. 50文章信息
- 郑仁华, 苏顺德, 肖晖, 许鲁平, 李林源, 林文龙, 张子文, 孟庆银
- Zheng Renhua, Su Shunde, Xiao Hui, Xu Luping, Li Linyuan, Lin Wenlong, Zhang Ziwen, Meng Qingyin
- 杉木优树多父本杂交子代测定及母本选择
- Female Selection Based on a Polycross Progeny Test of Cunninghamia lanceolata Plus Trees
- 林业科学, 2014, 50(9): 44-50
- Scientia Silvae Sinicae, 2014, 50(9): 44-50.
- DOI: 10.11707/j.1001-7488.20140906
-
文章历史
- 收稿日期:2013-11-04
- 修回日期:2014-03-16
-
作者相关文章


2. 福建省沙县官庄国有林场 沙县 365500;
3. 福建省三明市郊国有林场 三明 365000
2. Guanzhuang State-Owned Forest Farm of Fujian Shaxian 365500;
3. Sanming Suburban State-Owned Forest Farm of Fujian Sanming 365000
多父本杂交指用若干个父本的混合花粉对待测亲本授粉杂交的一种交配设计,虽然不能判定其子代父本,不宜作为选择群体开展进一步的选育,但较自由授粉对加性遗传方差和遗传增益的过高估算(Hallingbäck et al.,2013),多父本杂交对遗传参数的估算更具可靠性(王明庥,2001)。加之可操作性强,多父本杂交设计在林木育种中广泛应用,特别是对大量亲本进行同期评价时,效率高,相比其他交配方式具有明显优势(陈晓阳等,2005)。而在杉木(Cunninghamia lanceolata)亲本选择研究中,大多交配设计为自由授粉和全同胞交配设计(叶志宏等,1991;支济伟等,1994;阮梓材等,1994;李力等,2000b;翁玉榛,2008;何贵平等,2011),多父本杂交设计的报道较少(陈瑞生等,2011),对大量亲本进行同期多父本杂交子代测定的研究仅见李力等(2000a)关于12年生杉木分组群状多系杂交子代测定的报道。而对于未经一般配合力测定的亲本材料,一般配合力评选比特殊配合力评选更为重要(齐明,1996)。为了获得更为可靠的一般配合力测定信息,本文调查分析了1985年营建的杉木优树多父本杂交子代测定林27年生时的生长性状和木材基本性质,以期更为高效、准确地选择一批母本,充实育种材料。
1 材料与方法 1.1 试验材料及试验设计参试材料见李力等(2000a)报道。参试杂交组合137个,以福建省洋口国有林场杉木第1代种子园自由授粉混合种子为对照,共计138个处理。田间试验设计为随机区组设计,每个重复内群组随机排列,每个群组内杂交组合随机排列。重复6次,4株单列小区,造林密度为2 500株·hm-2。试验林保存完整,12年、27年林龄时保存率分别为91.3%和88.9%,小区株数调和平均数分别为3.63和3.51。
1.2 调查取样1996年12月,调查树高、胸径。2011年10月调查树高、胸径,同时选取1~5重复采集木芯样品。选取每个小区中生长最优株取样。测量胸径及胸径处4个方向的树皮厚度,随后在样木测胸径处上坡方位用直径为5 mm的生长锥取得通过髓心和树皮的一条完整无疵的木芯。现场测量木芯的全长、半长、红心材全长、红心材半长。木材的红心率=(红心长/去皮木芯长)2×100%;偏心率=︱半边木芯长/木芯长-0.5︱×100%;树皮率=100%-(D1.3-2×树皮厚)2/D1.32×100%。红心率、树皮率等百分率性状反正弦(arcsin $\sqrt x $)转换后统计分析。据树高和胸径估算单株立木材积(中华人民共和国农林部,1978): V=0.000 058 770 42D1.969983 1H0.896 461 57。以最大饱和含水量法测定木芯基本密度(Smith,1954): BG=1 / [(M-M1)/M1+1/DW],式中: BG为木芯基本密度(g·cm-3),M为木芯水饱和时的质量(g),M1为木芯烘干时(105 ℃)的恒质量(g),DW为构成细胞壁的木材物质的密度,取平均值为1.53。
1.3 统计分析方法按完全随机区组试验设计进行统计分析。对木材基本性质的小区平均值进行方差分析,线性模型为:${Y_{ij}} = \bar x + {B_i} + {F_i} + {E_{ij}}$,式中: Yij为第i个区组第j个家系的小区平均值,$\bar x$为群体平均值,Bi为第i个区组的效应值,Fj为第j个家系的效应值,Eij为机误。家系遗传力: $h_f^2 = r\delta _f^2/\left({\delta _e^2 + r\delta _f^2} \right)$式中: hf2家系遗传力,r为重复数,δf2为家系遗传方差,δe2为机误。对树高等生长性状的单株观测值进行方差分析,线性模型为:${Y_{ijk}} = \bar x + {B_i} + {F_j} + {\left({B \times F} \right)_{ij}} + {E_{ijk}}$,式中: Yijk为第i个区组第j个家系第k个观察值,$\bar x$为群体平均值,Bi为第i个区组的效应值,Fj为第j个家系的效应值,(B×F)ij为第i个区组第j个家系的效应值,Eijk为机误。家系遗传力:$h_f^2 = \delta _f^2/\left({\delta _e^2/nr + \delta _{rf}^2/r + \delta _f^2} \right)$式中: hf2为家系遗传力,δf2为家系遗传方差,n为小区调和平均株数,r为重复数,δrf2为区组、家系交互效应方差,δe2为机误。遗传增益:$\Delta G = S{h^2}/\bar x \times 100\% $,式中: ΔG为遗传增益,S为选择差,h2为遗传力,$\bar x$为对照或群体平均值(Wright,1981;孔繁浩,1985;Namkoong,1981;马育华,1982)。选择指数方程为:$I = \sum\limits_{i = 1}^n {{W_i}h_i^2{P_i}} $,式中: I为某杂交组合或单株聚合性状指数值,Wi为第i个性状权重(1/δ,δ为组合或单株均值标准差),hi2为第i个性状遗传力,Pi为第i个性状表型值(李光友等,2005)。
2 结果与分析 2.1 生长性状遗传变异表 1列出杉木优树多父本杂交子代测定林在单株水平上的生长表型分析结果。林龄12年时子代平均树高、胸径、单株立木材积分别为9.56 m,12.8 cm,0.073 686 m3,单株水平表型变异系数分别为18.49%,21.36%,52.68%;林龄27年时子代平均树高、胸径、单株立木材积分别为11.72 m,16.0 cm,0.141 820 m3,单株水平表型变异系数分别为18.65%,26.43%,66.85%。试验林生长中等,在单株水平上表型差异较大。
|
|
对试验林2个林龄段生长性状进行方差分析,并对差异显著的性状估算遗传参数 (表 2)。子代家系平均树高、胸径、单株立木材积在杂交组合间差异极显著。林龄12年时,子代家系平均树高、胸径和单株立木材积的遗传变异系数分别为2.14%,4.72%和10.94%,由遗传因素引起的立木材积变异可占群体均值的10.94%;3个性状的家系遗传力分别为0.143,0.468和0.406,杂交组合树高、胸径、立木材积的变异的14.3%,46.8%和40.6%可遗传至下一代。林龄27年时,子代家系平均树高方差分量估值约等于零,说明杂交组合间树高生长在表型上虽有差异,但由遗传因素引起的变异在总变异中分量极少,致使其方差分量估值近于零。可能是因为杉木达到成熟林龄时,树高生长进入缓慢期所致。子代家系平均胸径和立木材积的遗传变异系数分别为4.68%和7.33%,由遗传因素引起的立木材积变异占群体均值的7.33%;2个性状的家系遗传力分别为0.325和0.135,杂交组合胸径、单株立木材积的变异的32.5%和13.5%可遗传至下一代。
|
|
表 3列出了林龄27年时子代家系基本材性平均表现分析结果。优树多父本杂交子代测定林平均木材基本密度为0.330 35 g·cm-3。子代家系平均木材基本密度、红心率、偏心率和树皮率表型变异系数分别为9.92%,10.14%,48.40%和16.51%。而方差分析表明,杂交组合子代间的木材基本密度差异显著,红心率、偏心率、树皮率差异不显著,因此估算了木材基本密度的遗传参数(表 4)。子代家系平均木材基本密度的遗传变异系数、家系遗传力分别为2.52%和0.202,由遗传因素引起的木材基本密度变异可占群体均值的2.52%,木材基本密度的变异的20.2%可遗传至下一代。
|
|
|
|
兼顾生长、材性利用指数方程开展综合选择是林木多目标性状改良的高效方法(王润辉等,2012),而木材基本密度和木材力学性质相关性显著(成俊卿,1995)。已有研究表明,杉木木材基本密度与木材力学强度紧密相关,木材基本密度和生长性状独立遗传或呈负向弱遗传相关,为杉木生长、材性联合选择提供了可能(施季森等,1993;李晓储等,1999;王润辉等,2012)。前述遗传变异分析表明,林龄27年时,胸径遗传力较树高、立木材积高,木材基本密度受到一定程度的遗传控制。因此,以子代家系胸径、木材基本密度构建选择指数方程选择速生优质杂交母本。记选择指数为I,子代家系胸径均值的标准差为δD,胸径家系遗传力为hfD2,胸径表型值为D,子代家系木材基本密度均值的标准差为δρ,木材基本密度家系遗传力为hfρ2,木材基本密度表型值为ρ。构建选择指数模型:I=(hfD2·D)/δD+(hfρ2·ρ)/δρ。将试验林群体均值代入选择指数方程,得群体均值聚合指数值I0,再将各母本均值代入方程可得各母本聚合指数值,令其标准差为δI。指数值大于I0+δI的表现最好,是选择对象;指数值介于I0和I0+δI的表现较好,可作为潜力种质资源加以保存。
据上述方法,构建了速生优质母本选择指数方程:
$\begin{array}{l} I = 0.234\;88D + 11.329\;22\rho \\ {I_0} = 7.488\;9,{\delta _1} = 0.338\;55,{I_0} + {\delta _1} = 7.827\;5 \end{array}$
以母本指数值为据,筛选出‘F三明74-5’、‘F桃3’等11个速生优质杂交母本(表 5)。入选11个优良杂交母本子代林龄27年时,平均胸径、立木材积、木材基本密度分别为17.5 cm,0.174 251 m3和0.350 24 g·cm-3,分别比群体均值大9.38%,22.87%和6.02%,遗传增益均值分别为3.19%,3.14%和1.21%;指数值均值为8.082 5,比群体均值大7.93%。入选家系的未作为选择性状的其他木材基本性状表现也较好,如红心率均值为57.24%,比群体均值大5.41%;偏心率和树皮率均值分别为1.88%和10.65,分别比群体均值小10.48%和5.92%。
|
|
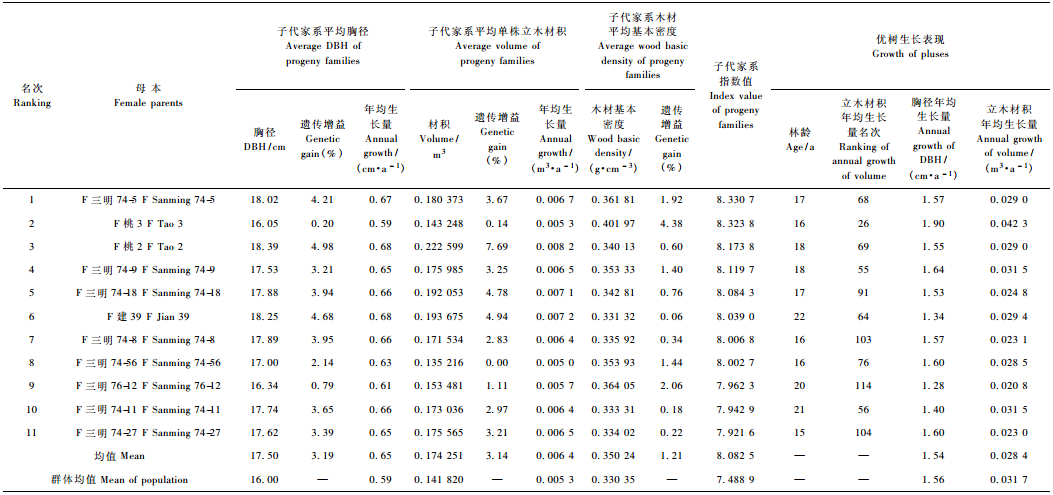
从表 5还可知,参试的137个母本的胸径、立木材积年均生长量总均值分别为1.56 cm·a-1和0.031 7 m3·a-1(优树选择时的调查数据),其多父本杂交子代分别为0.59 cm·a-1和0.005 3 m3·a-1。入选母本的优树胸径、立木材积年均生长量均值分别为1.54 cm·a-1和0.028 4 m3·a-1,其多父本杂交子代分别为0.65 cm·a-1和0.006 4 m3·a-1。优树年均生长优于其多父本杂交子代。这是因为优树为天然林中的自由授粉子代,授粉完全随机,而多父本杂交子代亲本间授粉不完全随机所致(李力等,2000a)。但应看到,多父本杂交子代测定重点在于测定母本的一般配合力,母本子代间在生长性状上的相对性较绝对性更具育种指导意义(王明庥,2001)。对比入选亲本优树及其子代排名可知,根据多父本子代测定,利用子代家系胸径和立木材积构建的指数方程筛选出的优良母本其优树在参试优树中年均生长排名基本在中后部。这一方面说明对优树进行遗传测定的重要性;另一方面也说明生长、材性联合选择与单纯的生长性状选择可能出现不同的选择结果,应根据育种目标采用适宜的选择方法。
2.4 入选率对中期选择效果的影响以试验林27年生时入选杂交组合为参照分析入选率对杉木优树杂交组合中期选择效果的影响。 当入选率介于1.00%~80.00%时,以2.00%为梯度设置不同入选率对杂交组合12年生和27年生时的胸径进行选择。同一入选率下,不同林龄具有相同的入选组合数,但入选参试组合不同。设定2个林龄段均入选组合的数量除以入选组合数,得12年生时的正确入选组合比率。因此可得每个入选率梯度相应的正确入选组合比率,以此作入选率-正确入选组合比率曲线图(图 1),分析入选率对杉木优树杂交组合中期生长性状选择效果的影响。
 |
图 1 入选率-正确入选组合比率曲线 Fig. 1 Selection ratio-correct selection ratio curve |
随着入选率增大,中期选择的正确入选组合比率增大。当入选率达79%时,正确入选组合比率可达85%以上。当入选率介于1%~19%时,增大入选率可有效提高中期选择的正确入选组合比率,每增大1%的入选率,可有效提高约3.7%的正确入选组合比率;当入选率介于19%~79%时,增大入选率对提高正确入选组合比率的作用较小,每增大1%的入选率,仅可提高约0.25%的正确入选组合比率。由于入选率与入选组合平均遗传增益成反比,因此,杉木杂交子代测定中期生长性状选择的入选率设置为19%时具有最高的选择效率,此时正确入选组合比率可达70%左右,在保证遗传增益的同时降低了中期选择的错选率和漏选率。
3 小结与讨论就生长性状而言,杉木优树多父本杂交子代测定林27年生时平均单株树高、胸径、立木材积分别为11.72 m,16.0 cm,0.141 820 m3,单株水平表型变异系数分别为18.65%,26.43%,66.85%。试验林生长中等,但表型差异较大。试验林12年生和27年生时,子代家系树高、胸径、立木材积在组合间差异均极显著,但胸径、立木材积受遗传因素影响较树高强烈,如27年生时树高方差分量估值约等于零,胸径和立木材积的家系遗传力分别为0.325和0.135。而木材基本性质分析结果表明,参试杂交组合子代间的木材基本密度差异显著,红心率、偏心率、树皮率差异不显著。子代家系木材平均基本密度的遗传变异系数、家系遗传力分别为2.52%和0.202。根据遗传变异分析结果,利用子代家系的胸径和木材基本密度构建选择指数方程对杂交母本进行了筛选。共计选出‘F三明74-5’、‘F桃3’、‘F桃2’等11个速生优质杂交母本,其多父本杂交子代27年生时平均胸径、立木材积、木材基本密度分别为17.5 cm,0.174 251 m3和0.350 24 g·cm-3,分别比群体均值大9.38%,22.87%和6.02%,遗传增益均值分别为3.19%,3.14%和1.21%。入选母本的优树与其多父本杂交子代的胸径和立木材积生长对比分析结果表明了进行优树遗传选择的重要性,也说明了在林木育种中,应根据育种目标采用适宜的选择方法。
入选的优良母本的子代平均胸径和立木材积遗传增益估值与李力等(2000a)的研究结果相近,但入选的母本或其排名却有区别。一方面是由于李力等(2000a)对优树的选择基于多点分析结果,而本研究基于单点分析。多点分析侧重点在于对参试材料进行多区域测试,其选择的材料可在各试验区域共同使用;而单点分析侧重于为特定区域选择优良材料。另一方面是由于选择林龄不同,优树的基因表达在时间上有差异所致;第三点是由于本研究利用胸径和木材基本密度构建选择指数方程对优树进行选择,而李力等(2000a)则利用树高、胸径、立木材积为指标进行选择。本研究中,对27年生时树高方差分量估值约等于零,胸径和立木材积的家系遗传力估值分别为0.325和0.135,较相关报道低。这主要是因为方差分量、遗传力估算受到立地环境、林龄、选择群体等多方面因素影响所致。特别是本研究试验林的立地为Ⅲ类立地,不利于喜肥沃、土层厚的杉木的生长,从而影响基于生长调查数据的方差分量估算结果。在林木育种中,对同一批材料进行评选是一种相对选择,这降低了参数估值不同而造成入选材料不同的风险性。
林木育种界认为,林龄越大,选择结果越可靠,多性状联合选择更有利于保持遗传多样性,就单点选择而言,本研究的选择结果具有可靠性,对优树亲本的利用具有指导意义。本研究对比分析试验林12年生时和27年生时对胸径的选择结果,认为杉木杂交子代测定中期生长性状选择的入选率为19%时具有最高的选择效率,此时正确入选组合比率可达70%左右,在保证遗传增益的同时降低了中期选择的错选率和漏选率。但这种对比分析仅基于试验林27年生时与12年生时的胸径对比,如果能与早、中期多年度数据进行比较将会取得更具说服力的结果。另外,由于交配方式为多父本杂交,试验林中的单株近亲率较高(母本不同,父本相同),故本研究未选择单株作为育种材料,但林分中的优良单株可作为无性繁殖材料利用。在今后的研究中,可利用分子标记技术对其进行亲本分析,发现其父本(El-Kassaby et al.,2009),将半同胞子代优良单株转化成全同胞子代单株利用,充分利用杂种优势,充实杉木育种资源,同时也可提高试验林的利用率。
| [1] |
成俊卿.1995.木材学.北京: 中国林业出版社,463-482.( 1) 1)
|
| [2] |
陈瑞生,石扬文,罗敬,等.2011.杉木多系授粉子代遗传变异及优良家系选择研究.种子,30(4): 54-57.(1)
|
| [3] |
陈晓阳,沈熙环.2005.林木育种学.北京: 高等教育出版社,134-135.(1)
|
| [4] |
何贵平,徐永勤,齐明,等.2011.杉木2代种子园子代主要经济性状遗传变异及单株选择.林业科学研究,24(1): 123-126.(1)
|
| [5] |
孔繁浩.1985.森林数量遗传学.南京: 南京林业大学遗传育种室,8.(1)
|
| [6] |
李光友,徐建民,陆钊华,等.2005.尾叶桉二代测定林家系的综合评选.林业科学研究,18(1): 57-61.(1)
|
| [7] |
李力,陈孝丑,曹汉洋,等.2000a.杉木分组群状多系杂交子代测定林的遗传分析.江西农业大学学报,22(3): 388-393.(5)
|
| [8] |
李力,施季森,陈孝丑,等.2000b.杉木两水平双列杂交亲本配合力分析.南京林业大学学报,24(5): 9-13.(1)
|
| [9] |
李晓储,黄利斌,王伟,等.1999.杉木木材基本密度变异的研究.林业科学研究,12(2): 179-184.(1)
|
| [10] |
马育华.1982.实验设计.北京: 农业出版社,241-260.(1)
|
| [11] |
齐明.1996.杉木育种中GCA与SCA的相对重要性.林业科学研究,9(5): 498-503.(1)
|
| [12] |
阮梓材,胡德活,陈仲.1994.杉木种子园建园亲本的综合选择.广东林业科技,(2): 14-19.(1)
|
| [13] |
施季森,叶志宏,翁玉榛,等.1993.杉木生长与材性联合遗传改良研究.南京林业大学学报,17(4):1-8.(1)
|
| [14] |
王明庥.2001. 林木遗传育种学.北京: 中国林业出版社,169.(2)
|
| [15] |
王润辉,胡德活,郑会全,等.2012.杉木无性系生长和材性变异及多性状指数选择.林业科学,48(3): 45-50.(2)
|
| [16] |
翁玉榛.2008.杉木第二代种子园自由授粉子代遗传变异及优良家系选择.南京林业大学学报: 自然科学版,32(1): 15-18.(1)
|
| [17] |
Wright J W. 1981.森林遗传学.郭锡昌,胡承海,译.北京: 中国林业出版社, 201-207.(1)
|
| [18] |
叶志宏,施季森,翁玉榛,等.1991.杉木十一个亲本双列交配遗传分析.林业科学研究,4(4): 380-385.(1)
|
| [19] |
支济伟,陈益泰,骆秀琴,等.1994.杉木主要材质性状配合力研究.林业科学研究,7(5): 531-536.(1)
|
| [20] |
中华人民共和国农林部.1978.LY208-77立木材积表.北京: 技术标准出版社.(1)
|
| [21] |
El-Kassaby Y A, Lstibůrek M.2009. Breeding without breeding.Genetics Research,91(2):111-120.(1)
|
| [22] |
Hallingbäck H R, Jansson G. 2013. Genetic information from progeny trials: a comparison between progenies generated by open pollination and by controlled cross. Tree Genetics & Genomes,9(3): 731-740.(1)
|
| [23] |
Namkoong G.1981.Introduction to quantitative genetics in forestry.London: Castle House Publications.(1)
|
| [24] |
Smith D M.1954. Maximum moisture content method for determining specific gravity of small wood samples. Forest Service, United States Department of Agriculture.(1)
|