 2014, Vol. 50
2014, Vol. 50文章信息
- 张俊艳, 成克武, 臧润国, 丁易
- Zhang Junyan, Cheng Kewu, Zang Runguo, Ding Yi
- 海南岛热带针-阔叶林交错区群落环境特征
- Environmental Characteristic Across a Tropical Coniferous-Broadleaved Forest Ecotone
- 林业科学, 2014, 50(8): 1-6
- Scientia Silvae Sinicae, 2014, 50(8): 1-6.
- DOI: 10.11707/j.1001-7488.20140801
-
文章历史
- 收稿日期:2013-04-24
- 修回日期:2013-06-14
-
作者相关文章



2. 河北农业大学园林与旅游学院 保定 071000
2. College of Landscape Architecture and Tourism, Agricultural University of Hebei Baoding 071000
生态交错区揭示了特定种及其生存环境在特定范围内的分布界限(Mayle et al., 2008),因此交错区被视为全球气候变化的敏感指示器(Wyckoff et al., 2010)。生态交错区生物多样性和生存环境复杂,是相邻生态系统镶嵌的复合体,受相邻生态系统相互作用的影响和调节,具有脆弱性和敏感性。目前,针叶林-草地生态交错区(Stott et al., 2003; Wyckoff et al., 2010)和高山林线生态交错区方面的研究较多(Jensen et al., 2008),而关于森林群落-森林群落生态交错区的研究较少(Goldblum et al., 2010),且多是关于北方落叶森林生态交错区(Elgersma et al., 2002; Bergeron et al., 2004)的研究。
针叶林主要分布在温带和寒带地区,在热带地区也有分布。我国热带针叶林以南亚松(Pinus latteri)为绝对优势种,仅混生少量阔叶树种,主要分布在海南霸王岭林区和琼中县。海南岛南亚松林与热带低地雨林和热带季雨林分布在同一海拔范围内(海拔<800 m),长期反复的刀耕火种和采割松脂及多次大规模商业采伐,使南亚松林受到了严重破坏(丁易等,2011a)。当这3种低海拔的热带林受到干扰破坏后,常常会形成针叶林与阔叶林的交错区。与自然形成的生态交错区不同的是,南亚松林针阔交错区(简称针阔交错区)在一定程度上受到人为干扰的影响,但是一种稳定性的群落类型。目前我国热带针叶林的研究只关注南亚松的林分结构和分布等方面(臧润国等,2010),关于热带针叶-阔叶林交错区林分状况的研究很少,更没有针对针叶-阔叶林交错区林分环境特征方面的研究。本研究采用样带法研究海南岛霸王岭林区典型的热带针叶林-阔叶林交错区的环境特征,探讨环境因子随群落类型的变化规律,为今后更系统、深入地研究针叶-阔叶林交错区的群落结构、多样性、动态和功能机制等奠定基础。
1 研究区概况海南霸王岭国家级自然保护区位于海南省西南部昌江县和白沙县境内(108°58′—109°53′E, 18°53′—19°20′N),总面积7.1万hm2,海拔50~1 654 m。该地区属热带季风气候,干湿季明显。年均气温24.2 ℃,年均降水量1 559~2 447 mm, 年蒸发量983~1 313 mm, 空气相对湿度70%~80%,5—10月为雨季,11月至翌年4月为旱季。海南岛霸王岭林区的南亚松林是我国面积最大的热带天然针叶林。南亚松林土壤多为砂壤,含石砾,土壤结构疏松,土层较深厚,一般达100~150 cm, 山腰以上在100 cm以下,山脊不足50 cm。植物根多分布在50~80 cm的土层。南亚松林林冠层以南亚松占绝对优势,最高可达40 m, 胸径最大可达2 m, 在天然南亚松群落中南亚松多为该群落的优势种,有些地区伴有青梅(Vatica mangachapoi)、毛叶青冈(Cyclobalanopsis kerrii)、黄杞(Engelhardtia roxburghiana)、银柴(Aporusa dioica)、余甘子(Phyllanthus emblica)和烟斗柯(Lithocarpus corneus)等。草本层植物种类以禾本科植物居多,如华须芒草(Andropogon chinensis)、短梗苞茅(Hyparrhenia dipl and ra)和红裂稃草(Schizachyrium sanguineum)等(蒋有绪等,2002)。针阔交错区的阔叶树种以热带季雨林种为主,个体数较多的科为大戟科(Euphorbiaceae),其次为茜草科(Rubiaceae),常见种有银柴、厚皮树(Lannea corom and clica)和黄牛木(Cratoxylum ligustrinum)等。阔叶林区的树种有银柴、黄牛木、光叶巴豆(Croton laevigatus)和山柑算盘子(Glochidion sphaerogynum)等,多数是刀耕火种弃耕地恢复早期种,还有黄杞、木荷(Schima superba)和青梅等热带低地雨林常见优势种(丁易等,2011b)。
2 研究方法 2.1 样地设置与调查2012年2—6月,对海南岛霸王岭林区的南亚松分布区进行了全面调查。在针叶林与阔叶林交错的地点布设13条典型样带,样带从典型针叶林区开始,横穿针阔交错区到典型的阔叶林区为止,长90~160 m, 宽20~60 m。在每条样带的3种群落类型内各随机设置6块10 m×10 m样地,对样地内所有胸径>1 cm的乔、灌木树种进行每木检尺,详细记录树种的名称、树高、胸径,并记录样地坡度、枯枝落叶层厚度。其中,样带1和10针叶林区位于山体下坡,阔叶林区位于山体上坡,平均坡度小于5°;样带2~9和样带11~13针叶林区位于山体上坡,阔叶林区位于山体下坡,坡度5~10°。样带1和3为废弃围猎场恢复起来的次生林,样带2和4~13为少数民族刀耕火种弃耕地恢复起来的次生林。调查区内林冠层全部为南亚松,阔叶树种很少能够进入到林冠层。3种群落类型的林分因子差异见表 1。
|
|
在每块样地的中心,使用英国Hemiview林冠分析仪(HMV1v8,Delta-T Devices Ltd, Cambridge, UK)测定林冠开阔度(%)。同时,在每块样地的四角和中心位置分别测量林下凋落物层厚度,求其平均值得到林分凋落物层厚度(LD, cm)。
2.2 土壤样品采集及分析采用土钻法采集土样。在每块10 m×10 m样地四角及中心位置收集表层土壤(0~30 cm),均匀混合为1个土样,即每条样带每种群落类型6个土壤样品,共得到234份土壤混合样。每份土壤混合样取适量部分放入铝盒中带回实验室称鲜质量,土壤样品在105 ℃的烘箱中烘干至恒质量后称干质量,计算土壤含水量(%),另一部分利用土壤袋收集200 g带回室内自然风干,然后采用重铬酸钾外加热容量法测定土壤有机质含量;采用电位法测定土壤pH;分别用凯氏定氮法、NaOH熔融-火焰光度法和钼蓝光度法测定全氮、全磷和全钾含量;分别用碱解扩散法、钼蓝光度法和乙酸铵提取火焰光度法测定有效氮、有效磷和有效钾含量;用烘干法测定土壤砂砾含量。所有测定方法均依据国家林业行业标准执行(张万儒等,1999)。
2.3 数据处理采用方差分析和多重比较分析13条样带3种群落类型15个环境因子的差异。对每条样带每种群落类型中的15个环境因子数据分别取算术平均值,从而得到39 × 15的群落类型与环境因子矩阵,采用Pearson相关分析法对该矩阵进行环境因子相关性分析,相关性大小用Student’s t 检验,并进行主成分分析。本研究所有数据统计分析均在R-2.15.0程序中进行。
3 结果与分析 3.1 环境因子特征3种群落类型的林冠开阔度、直射光透过率、散射光透过率和总透光率均表现为针叶林区>针阔交错区>阔叶林区,针叶林区和针阔交错区显著大于阔叶林区,针叶林区和针阔交错区差异不显著。3种群落类型凋落物厚度整体差异显著,表现为针叶林区>针阔交错区>阔叶林区,针阔交错区与阔叶林区无显著差异。土壤含水量表现为阔叶林区>针阔交错区>针叶林区,但差异不显著。土壤有机质含量表现为阔叶林区>针阔交错区>针叶林区,针叶林区和阔叶林区之间差异显著,针阔交错区均无显著差异。土壤砂砾含量在3种群落类型之间无显著差异,表现为针叶林区>针阔交错区>阔叶林区。土壤pH表现为阔叶林区>针阔交错区>针叶林区的趋势,交错区和针叶林区无显著差异。全钾含量表现为针叶林区>针阔交错区>阔叶林区,针叶林区显著大于阔叶林区和针阔交错区;全氮、有效氮、有效磷和有效钾在针叶林区和阔叶林区之间均存在显著差异,均表现为阔叶林区>针阔交错区>针叶林区。
3.2 环境因子相关性分析3种群落类型环境因子相关性见表 2,林冠开阔度、直射光透过率、散射光透过率和总透光率4个因子相互间均极显著相关(P< 0.001),同时这4个因子与土壤砂砾含量极显著相关(P < 0.01),与土壤pH和土壤有机质含量显著相关(P < 0.05)。凋落物厚度与全钾含量极显著相关(P < 0.01),与其他因子之间无显著相关(P>0.05)。土壤含水量与其他因子之间无显著相关(P>0.05)。土壤有机质含量与全氮含量、有效氮含量和有效钾含量的相关性最大(P < 0.001);全磷含量与其他环境因子之间均无显著相关性(P>0.05)。土壤pH与全钾含量、有效氮含量和有效钾含量极显著相关(P < 0.01),与4个光因子(林冠开阔度、直射光透过率、散射光透过率和总透光率)、土壤砂砾含量、土壤有机质/全氮显著相关(P < 0.05),与全磷含量、凋落物厚度相关性不显著(P>0.05)。全氮含量与有效氮含量和有效钾含量的相关性较高(P < 0.01),全钾含量与有效钾含量相关性较小(P>0.05),全磷含量与有效磷含量相关性最小(P>0.05)。全氮含量、有效氮含量和有效钾含量与多数因子相关性都显著,土壤含水量和全磷含量与多数因子相关性都不显著。
|
|
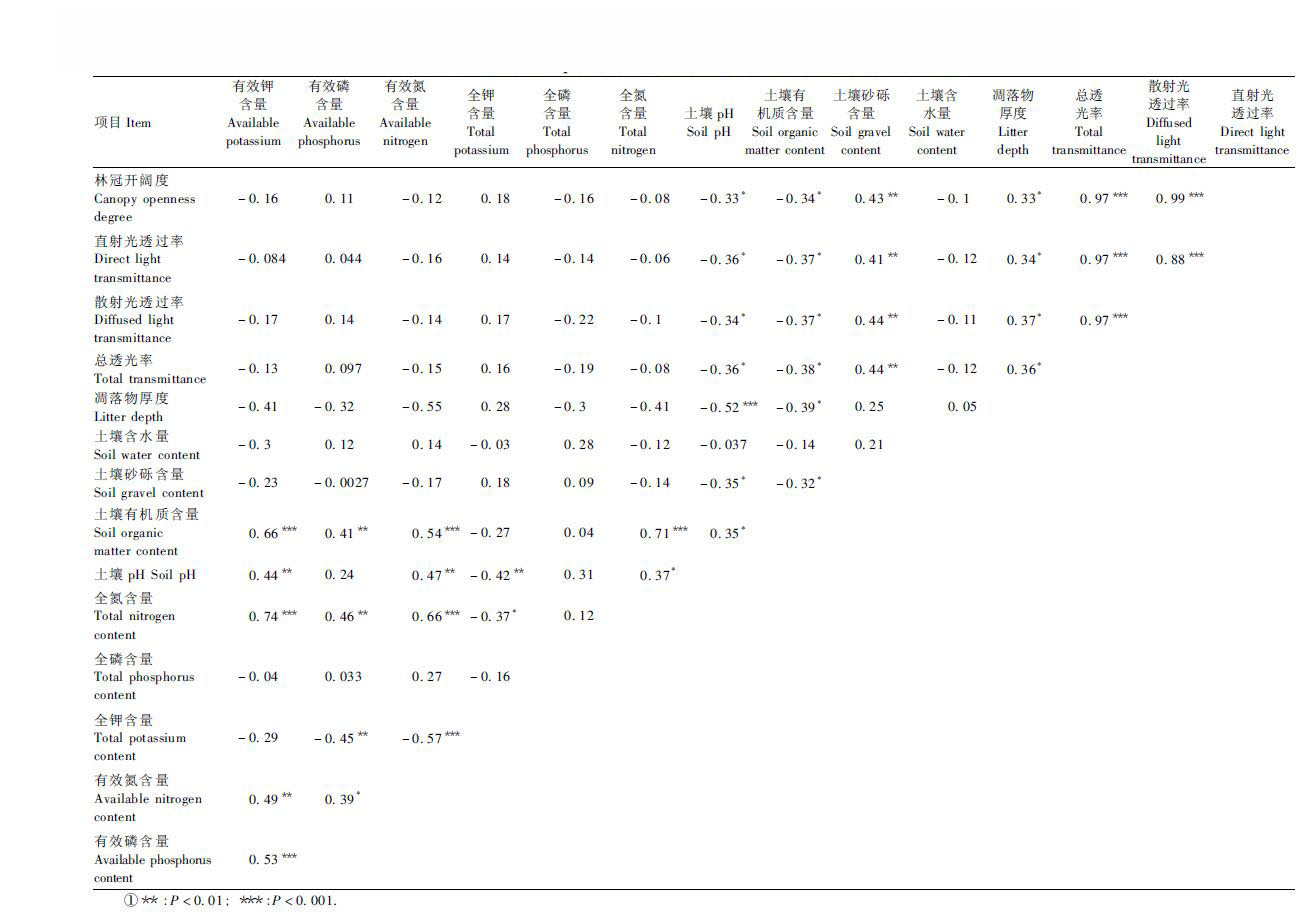
3种群落类型环境因子的主成分分析(principal correspondence analysis, PCA)结果见表 3。PCA前2轴的累积解释方差比例为57.7%,前4轴的累积解释方差比例达75.0%。环境因子在前2个排序轴的负荷值较高,所以根据前2个PCA轴分析环境因子的作用大小。第1轴主要反映土壤pH、土壤有机质含量、凋落物厚度、林冠开阔度、直射光透过率、散射光透过率、总透光率和有效钾含量的变化;第2轴主要反映全氮含量和有效磷含量的变化。
|
|
3种群落类型随环境因子的变化呈现出明显的变化趋势(图 1)。针叶林区和阔叶林区分布在PCA排序轴的两边,针阔交错区介于针叶林区和阔叶林区之间。沿着PCA第1排序轴从左到右,随着土壤pH降低,凋落物厚度减少,土壤光环境增强,林分由阔叶林区到交错区再到针叶林区过渡;沿着PCA第2排序轴从下到上,随着土壤全氮含量和有效磷含量增加,林分由针叶林区到交错区再到阔叶林区过渡。

|
图1 3种群落类型随环境因子变化的PCA排序 Fig.1 Biplot of principal correspondence analysis for the three community types along environmental factors |
3种群落类型光因子特征均表现为针叶林区>针阔交错区>阔叶林区。这是因为针叶林区林分密度小、植株较为高大、林冠层稀疏,导致林冠开阔度最大;阔叶林区林分密度最大,林冠开阔度最小;而在针阔交错区,虽然林分密度相对针叶林区有所增加,但由于林分内植株个体高度差异相对较大、物种更替速率较快,导致林冠开阔度仍大于阔叶林区。森林群落中光资源具有高度的时间和空间异质性,不同光条件下生存的种具有不同的生理和形态特征(Ameztegui et al., 2011)。针叶林区南亚松树种、针阔交错区的季雨林落叶树种及阔叶林区的热带低地雨林种对光的利用程度也与林分内的光照强度相一致。凋落物是森林生态系统的重要组分,3种群落类型凋落物厚度表现为针叶林区>针阔交错区>阔叶林区。这是因为针叶林区林下干燥且除少量季雨林种外全为南亚松种,林下凋落物多是松树的针叶、枯枝、皮及成熟后凋落下的果壳,不易腐烂分解;而阔叶林区凋落物多为热带低地雨林种及一部分热带季雨林种,加上林下潮湿、土壤肥力较大,导致凋落物分解速度较快。
土壤有机质含量在针叶林区和针阔交错区间无显著差异,但都显著低于阔叶林区。这是由于针叶林区和针阔交错区多位于山坡的中上部,保水性差,加上强烈光照的影响,土壤较为干旱,因此土壤含水量和土壤肥力都最差;而阔叶林区林分密度和物种丰富度都大于针阔交错区和针叶林区,动植物残体种类较多,为土壤提供了较多的有机质源(Kammer et al., 2009),可有效减少水分蒸发,因此土壤肥力和土壤含水量都较高。本研究得到,土壤pH表现为阔叶林区>针阔交错区>针叶林区,但针阔交错区与针叶林区无显著差异、与阔叶林区显著差异,这可能与南亚松的存在有关。随着林分由针叶林经针阔交错区到阔叶林的转变,土壤全氮、有效氮、速效磷和速效钾含量均表现出阔叶林区>针阔交错区>针叶林区,这也可能与林分的物种组成及其凋落物特性对土壤的长期作用有关(夏汉平等,1997)。长期反复刀耕火种也影响森林群落土壤砂砾含量的变化(Chazdon, 2003)。
从相关性分析(表 2)可以看出,土壤pH与有效氮含量的相关性最大。Koptsik等(2003)研究认为林冠开阔度影响森林土壤的pH, 同时森林土壤的理化性质,尤其是土壤pH和速效养分也受气温、降雨、凋落物归还量以及土壤微生物的数量和种类等其他环境因子的影响。本研究表明,土壤含水量与土壤有效钾含量和总氮含量的相关性最大,但与其他因子表现出较小的相关性。在一定含量范围内,土壤有机质含量与土壤肥力水平(如土壤氮、土壤磷、土壤有效氮)呈正相关。磷由于易被土壤、微生物和凋落物所固定,不易被活化和淋失,因此土壤全磷含量与其他因子相关性较小。
目前国内尚无关于热带针叶林-阔叶林交错区的文献,国外很多以热带山地森林为对象的研究都认为海拔、地形、干扰及物种的不同导致不同森林类型间存在边界(Ashton, 2003; Martin et al., 2007)。其中,热带针叶林-云雾林交错区的研究认为,随着林分由云雾林到针叶林的转变,林分湿度和温度逐渐降低,针叶林最为干旱,火干扰较为频繁(Martin et al., 2011)。虽然与本研究的研究区域不同,但交错区的环境特征变化趋势表现出一致性。
凋落物分解速率在雨季和旱季不同,且分解速率与环境因子密切相关(张德强等,2000)。南亚松林中凋落物层较厚、分解周期较长,本研究只分析了凋落物层的厚度,未考虑凋落物分解特性对不同林分区域环境因子的关系,因此对热带针-阔叶林交错区群落环境特征还需进一步研究。
| [1] |
丁易, 臧润国. 2011a. 采伐方式对海南岛霸王岭热带山地雨林恢复的影响. 林业科学, 47(11): 1-5.( 1) 1)
|
| [2] |
丁易, 臧润国. 2011b. 海南岛霸王岭热带低地雨林植被恢复动态. 植物生态学报, 35(5): 577-586.(1)
|
| [3] |
蒋有绪, 王伯荪, 臧润国, 等. 2002. 海南岛热带林生物多样性及其形成机制. 北京: 科学出版社.(1)
|
| [4] |
夏汉平, 余清发, 张德强. 1997. 鼎湖山3种不同林型下的土壤酸度和养分含量差异及其季节动态变化特性. 生态学报, 17(6): 645-653.(1)
|
| [5] |
臧润国, 丁易, 张志东, 等. 2010. 海南岛热带天然林主要功能群保护与恢复的生态学基础. 北京: 科学出版社.(1)
|
| [6] |
张德强, 叶万辉, 余清发, 等. 2000. 鼎湖山演替系列中代表性森林凋落物研究. 生态学报, 20(6): 938-944.(1)
|
| [7] |
张万儒, 杨光澄, 屠星南. 1999. 森林土壤分析方法. 北京: 中国标准出版社.(1)
|
| [8] |
Ameztegui A, Coll L. 2011. Tree dynamics and co-existence in the montane-sub-alpine ecotone: the role of different light-induced strategies. Journal of Vegetation Science, 22(6): 1049-1061.(1)
|
| [9] |
Ashton P S. 2003. Floristic zonation of tree communities on wet tropical mountains revisited. Perspectives in Plant Ecology, Evolution and Systematics, 6(1): 87-104.(1)
|
| [10] |
Bergeron Y, Gauthier S, Flannigan M, et al. 2004. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern quebec. Ecology, 85(7): 1916-1932.(1)
|
| [11] |
Chazdon R L. 2003. Tropical forest recovery: legacies of human impact and natural disturbances. Perspectives in Plant Ecology, Evolution and Systematics, 6(1): 51-71.(1)
|
| [12] |
Elgersma A M, Dhillion S S. 2002. Geographical variability of relationships between forest communities and soil nutrients along a temperature-fertility gradient in norway. Forest Ecology and Management, 158(1): 155-168.(1)
|
| [13] |
Goldblum D, Rigg L S. 2010. The deciduous forest-boreal forest ecotone. Geography Compass, 4(7): 701-717.(1)
|
| [14] |
Jensen C, Vorren K D. 2008. Holocene vegetation and climate dynamics of the boreal alpine ecotone of northwestern fennoscandia. Journal of Quaternary Science, 23(8): 719-743.(1)
|
| [15] |
Kammer A, Hagedorn F, Shevchenko I, et al. 2009. Treeline shifts in the ural mountains affect soil organic matter dynamics. Global Change Biology, 15(6): 1570-1583.(1)
|
| [16] |
Koptsik S, Koptsik G, Livantsova S Y, et al. 2003. Analysis of the relationship between soil and vegetation in forest biogeocenoses by the principal component method. Russian Journal of Ecology, 34(1): 34-42.(1)
|
| [17] |
Martin P H, Fahey T J, Sherman R E. 2011. Vegetation zonation in a neotropical montane forest: environment, disturbance and ecotones. Biotropica, 43(5): 533-543.(1)
|
| [18] |
Martin P H, Sherman R E, Fahey T J. 2007. Tropical montane forest ecotones: climate gradients, natural disturbance, and vegetation zonation in the cordillera central, dominican republic. Journal of Biogeography, 34(10): 1792-1806.(1)
|
| [19] |
Mayle F E, Power M J. 2008. Impact of a drier early-mid-holocene climate upon amazonian forests. Philosophical Transactions of the Royal Society B: Biological Sciences, 363(1498): 1829-1838.(1)
|
| [20] |
Stott P, Mast J N, Veblen T T, et al. 2003. Disturbance and climatic influences on age structure of ponderosa pine at the pine/grassland ecotone, colorado front range. Journal of Biogeography, 25(4): 743-755.(1)
|
| [21] |
Wyckoff P H, Bowers R. 2010. Response of the prairie-forest border to climate change: impacts of increasing drought may be mitigated by increasing CO2. Journal of Ecology, 98(1): 197-208.(2)
|