 2013, Vol. 49
2013, Vol. 49文章信息
- 陈朋, 于雪丹, 张川红, 郑勇奇, 孙圣, 程蓓蓓, 朱从波
- Chen Peng, Yu Xuedan, Zhang Chuanhong, Zheng Yongqi, Shun Sheng, Cheng Beibei, Zhu Congbo
- 中国特有种血皮槭的天然更新
- Natural Regeneration of Acer griseum, an Endemic Species in China
- 林业科学, 2013, 49(3): 159-164
- Scientia Silvae Sinicae, 2013, 49(3): 159-164.
- DOI: 10.11707/j.1001-7488.20130322
-
文章历史
- 收稿日期:2012-03-08
- 修回日期:2012-12-27
-
作者相关文章



2. 南京林业大学风景园林学院 南京 210037;
3. 河南宝天曼国家级自然保护区管理局 内乡 474350
2. College of Landscape Architecture, Nanjing Forestry University Nanjing 210037;
3. Henan Baotianman National Nature Reserve Authority Neixiang 474350
血皮槭(Acer griseum),又名马梨光,槭树科(Aceraceae),槭树属(Acer),落叶乔木,高达20m,为中国特有种、濒危种(汪松等,2004)。血皮槭夏季树干树皮有卷曲状剥落,树皮色彩奇特,呈鲜明的橘褐色或红褐色,尤其在冬季,观赏价值极高。其树叶春、夏季为绿色,叶脉、叶柄及新梢为红色。血皮槭秋季10,11月叶变色,从黄色、橘黄色至鲜红色,落叶晚,非常美观;且树冠较大,叶多而密,遮荫良好,是槭树类中经济价值较高的绿化树种,宜引种为行道树或庭园树种,常作为庭园主景树(钱又宇等,2008)。血皮槭树干挺直,木材坚硬,材质细密,可作车轮、家具、农具、枕木及建筑材料;纹理清晰美观可用以制造乐器和工艺品;其树皮纤维可为造纸及人造棉提供原料,还可提取药物(中国植物志编辑委员会,1981)。血皮槭主要集中分布在我国中部地区,包括湖南、湖北、重庆、河南、山西、陕西和甘肃等省市,亚热带、北亚热带和暖温带等3个气候带。生长于海拔1500~2000m疏林中(中国植物志编辑委员会,1981)。
在《中国物种红色目录》(汪松等,2004)中记载血皮槭的过去3个世代种群数量至少减少50%,但濒危原因目前尚未有文献报道。血皮槭虽然观赏价值很高,由于其繁殖很困难,除北京植物园少量种植外,在国内园林绿化中很少见。早在1901年前英国便从我国引种(托比·马斯格雷夫等,2005),栽植于世界著名的植物园或苗圃或公园内,但没有大面积种植。目前,对于此树种国内外研究报道较少,国外研究大多集中于繁殖,包括种子繁殖和无性繁殖(张川红等,2008)。国内杜丽雁(2005)曾对血皮槭种子育苗做过报道,陈丽等(2010;2012)开展了血皮槭的组培研究,李倩中等(2010)以槭属植物为研究对象,通过SRAP标记在分子水平上探讨了31种槭树的亲缘关系,将它们分成6大类:其中血皮槭与三角槭(Acer buergerianum)、樟叶槭(Acer cinnamomifolium)、红翅槭(Acer fabri)分为一组,亲缘关系较近。而对于血皮槭野生资源的生境、天然更新及生物学特性等研究几乎空白,从而导致血皮槭的价值也没有得到充分开发和利用。本研究将从血皮槭的生境调查、土壤种子库及其天然更新进行系统研究,了解天然群体的生境与更新状况,为其濒危机制的研究提供理论依据。
1 研究方法 1.1 文献查阅及标本核对查阅《中国树木志》及地方植物志、树木志和中国科学院植物所标本馆血皮槭的标本,判断该树种的分布范围。
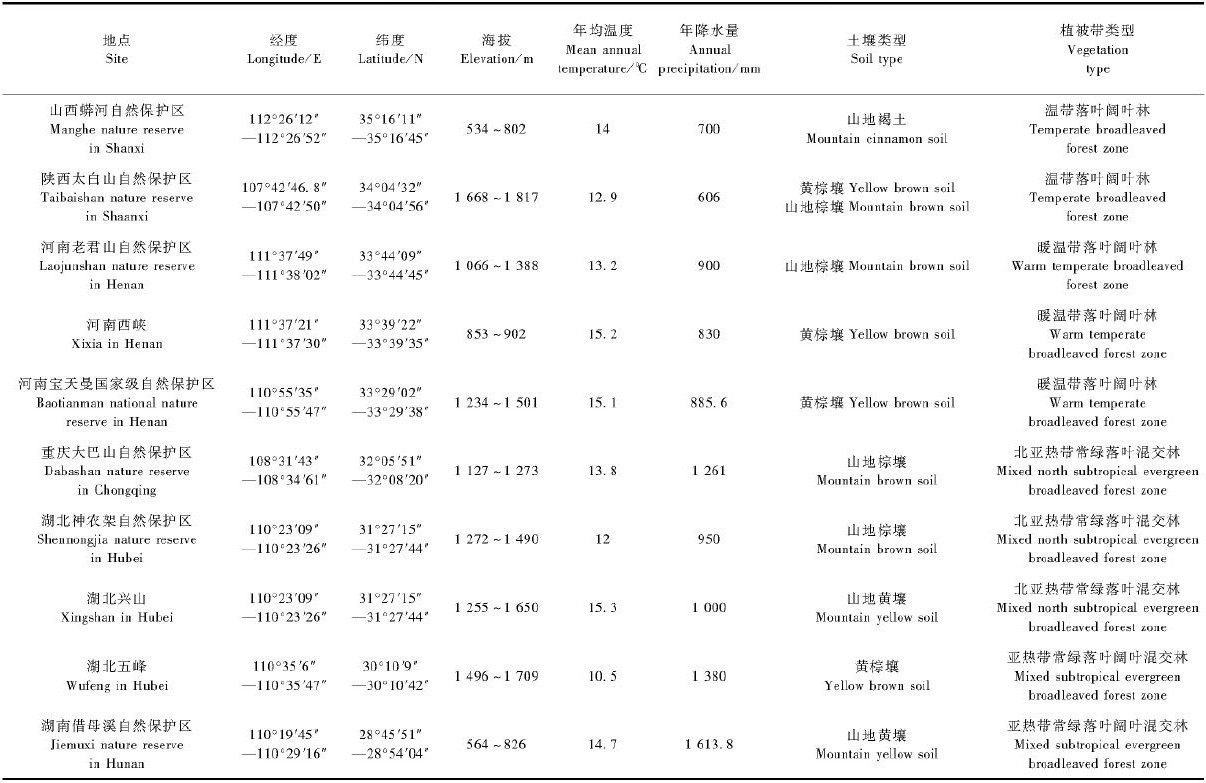
1.2 实地调查1)调查地点概况 根据文献和标本研究结果,确定在血皮槭的分布范围内选取山西蟒河自然保护区,陕西太白山自然保护区,河南宝天曼国家级自然保护区、老君山自然保护区、西峡,湖北的五峰、兴山、神农架林区,湖南借母溪自然保护区,重庆大巴山自然保护区等10个分布地点,采用线路调查法(彭东辉等,2006)进行调查,调查地点的经纬度、气候因子、海拔、坡向、土壤类型、群落类型等见表 1,并对原生境和植株生长状况进行调查、拍照和GPS卫星定位。
|
|
2)天然更新调查 选取血皮槭分布区的5个典型地点(重庆大巴山、陕西太白山、山西蟒河、河南老君山、湖南借母溪)进行天然更新调查。调查采用张川红等(2005)的方法,按海拔从低到高选择单株,测定单株的树高和胸径(或地径),观察其周围是否存在幼树(或幼苗)。如果有更新苗,以其为圆心,沿下风向设置取一个45°扇形样地,扇形半径30m,通过刨根挖出其根系,判断更新苗是实生苗、根蘖苗或桩蘖苗。每个地点选择30株母株,调查更新苗离母株的距离(m),幼苗的高度(cm)、地径(cm)及光合有效辐射。
3)土壤种子库调查 于2011年5月至10月在重庆大巴山(8株)、陕西太白山(7株)、山西蟒河(5株)、河南老君山(6株)、湖南借母溪(6株),从水平和垂直2个方向调查血皮槭的土壤种子库,母树间距大于50m。由于种子在土壤中的分布是极不均匀的,因此减少取样的随机误差,提高取样的精确性,在选定的血皮槭样株周围,在上风向、下风向、垂直风向按树高0.5,1,2倍距离取样,以“品”字形排列设置3个大样方,在大样方内在随机取3个20cm×20cm小样方,每个小样方分为枯枝落叶层,0~10cm,10~20cm 3个土层取土壤(许建伟等,2010),用筛网过滤后种子装袋保存,室内查数其中的血皮槭种子数量,同时依据不同深度做方差显著性检验,并检查种子的完整性和空壳情况并统计分析其空种率(付增娟等,2006)。
2 结果与分析 2.1 血皮槭分布的生境特点文献研究与实地调查发现,血皮槭是亚顶级群落植物,散生分布在山地阔叶混交林、山地针阔叶混交林、山地山顶灌丛3种植被带类型中。在这些群落中,除了在山顶灌丛群落中作为上层树种外,在其他群落中均是伴生树种,处于群落结构的第2层。血皮槭属于中、高山植物,除了在河南西峡、老君山和湖南借母溪在1000m以下有分布外,主要集中分布在1000~1800m。
调查发现,血皮槭自然分布区以山地为主,而且全部分布在半阳坡、半阴坡、阴坡以及沟谷环境中,未发现在阳坡分布(表 1),表明血皮槭喜阴湿环境,空气湿度也影响其自然分布。土壤类型以山地棕壤、黄棕壤、山地褐土为主,表明其喜微酸性土壤;土层厚度从裸露的岩石到30cm左右,大多在10cm左右,而且部分甚至生长在非常贫瘠的土层或者岩石缝隙中,说明血皮槭具有较强的耐瘠薄能力。
调查发现,野生血皮槭的胸径范围为10~30cm,平均25cm左右。最大的血皮槭在河南老君山内发现,其树高14m,地径63.5cm。野生血皮槭平均树高在8.5~12m。没有发现特别高大的血皮槭单株。野生血皮槭的成熟单株存在不结实的现象,在重庆大巴山、河南宝天曼和陕西太白山的血皮槭未见到结实单株(表 2)。而其他调查地点的结实大树也不多,统计表明接近80%的大树不结实。调查还发现,只有生长在林缘或空旷地或林窗的血皮槭才大量结实。
|
|
通过调查分析,种子水平分布很不均匀,离母株越远种子越少。在调查的5个地点中,在离母树0.5倍距离种子库中种子数量最多,在0.5,1,2倍树高距离的平均种子数量分别是460,208,68粒·m-2,种子在土壤中的数量与离母树的距离成负相关,距离越远种子越少(图 1)。同时,土壤种子库的水平分布与风向有关系,顺风向有助于种子传播。在5个调查地点中顺风向种子数量分别为65~142粒·m-2,相比于垂直风向为38~87粒·m-2,顺风向种子数量明显较多,所以在顺风向种子传播距离较远。风是种子传播的主要媒介之一。

|
图1 5 个调查地点土壤种子库水平方向种子分布 Fig.1 Seed horizontal distribution in soil seed bank |
在垂直方向的3个土层中,在枯落层中种子分布最多,其中最多的是湖南借母溪有172粒·m-2,最少的是河南老君山56粒·m-2;种子在0~10cm土层和大于10cm的土层中的数量分布明显比枯落层少,最多的湖南借母溪在0~10cm土层只有42粒·m-2,是相应枯落层的四分之一(表 3)。
|
|
土壤库中种子的空种率极高,5个地点中空种率最高达81%,最低也有43%,这也是天然更新植株较少的原因之一。
1)种子繁殖更新及实生苗生长状况5个观测地点中都发现有实生更新苗,30株母树中周围能发现有实生更新苗的母树数量在4~8株,在每株母树周围更新苗数量大约4株左右,在湖南借母溪发现更新苗数量最多,有28株更新苗,而且长势最好(表 3);山西蟒河和陕西太白山的更新苗数量较少,且长势一般;调查发现,在重庆大巴山和湖南借母溪幼苗的光合有效辐射比较高,陕西太白山和山西蟒河的实生更新苗光合有效辐射比较低,因此推测,幼苗的生长状况受生境光照的影响较大。
在5个调查地点中,只有在重庆大巴山和湖南借母溪有发现根蘖苗,分别占总数的10.53%和3.57%(表 4),根蘖苗基本都是在离母树比较近的裸露根系中萌发出来,没有发现从深层土壤根系中萌发的根蘖苗。除了在陕西太白山和重庆大巴山没有发现桩蘖苗外,其余3个调查地点均发现桩蘖苗,但是数量不多,在山西蟒河、河南老君山和湖南借母溪,桩蘖苗更新数分别占调查株数的42%,3%和28%,这表明血皮槭天然更新以实生更新为主。
|
|
2)天然更新与土壤种子库的关系在重庆大巴山、陕西太白山、山西蟒河、河南老君山、湖南借母溪5个调查地中,在周围发现实生苗的母树,同时随机选取相同数量没有实生苗的单株作土壤种子库调查。经调查发现,在以上5个调查地中,分别有8,7,5,6,6株母树周围土壤库中有种子发现,在有实生苗的母树周围土壤库中均发现种子,只有在陕西太白山、山西蟒河、湖南借母溪各发现1株在没有实生苗的母树周围土壤库中有种子。结果表明有些母树周围的土壤库中即使有种子,但没有幼苗更新,这说明种子在自然界的萌发有一定的难度。另外,还有一部分母树周围没有更新苗的原因之一是土壤种子库中没有种子。土壤种子库是植物繁殖体长期贮存的天然库,推测导致此结果的原因是野生群体中有些血皮槭大树因为某些原因导致不结实,因此其周围土壤库中没有种子。
3 结论与讨论调查结果表明:野生血皮槭单株结实量极低或不结实,结实种子空种率极高。除了在山地顶灌丛群落中为上层树种,在山地阔叶混交林和山地针阔叶混交林群落中,均处在群落结构的第2层,林分郁闭度基本都处于0.6以上,这决定了血皮槭生境的光照条件较差。光照不足时,植物的成花过程将受到显著抑制(王沙生等,1991),所以生长在空旷地的树木开花结实比林木早且多。因此,血皮槭作为伴生树种,生境的光照强度严重不足,导致结实量低或不结实。因此推测较差的光照条件是血皮槭结实率低的主要原因之一。
土壤种子库是植物天然更新与植被恢复的物质基础(Moles et al., 1999),可以揭示种群和群落动态。森林土壤种子库的密度一般在100~1000粒·m-2(Halpern et al., 1999),但调查结果表明:血皮槭土壤种子库与其他植物相比数量要少一些,最多的湖南借母溪为172粒·m-2,最少的河南老君山仅有52粒·m-2,其密度较低。血皮槭结实量少导致土壤种子库内种子数量少,除此之外,还与血皮槭生长环境有关。血皮槭大部分生长在陡峭的山坡或者岩石缝隙中,致使种子散落后落入沟底随水流走,不易在地表及土壤内存留,所以土壤种子库数量较少,而且分布不均。但湖南借母溪的血皮槭枯落物层较厚,有利于种子散落后存留,表层土壤深厚疏松,有利于种子从枯落物中进入表层土中,所以相对于其他调查地点种子库中种子数量稍多。
本研究结果表明血皮槭主要以种子繁殖为主,并且更新的数量不多。在调查的5个地点中都发现有实生天然更新苗,但是数量不多,最多的是湖南借母溪,5株母树周围样地内发现26株实生更新苗。在血皮槭分布的山坡下缘沟壑或者坡地以及平坦的山地中几乎也很难发现有血皮槭更新苗出现,这也许与空种率高、种子萌发困难、种子被动物吞食有关。血皮槭野生单株种子空种率60%左右,种子库内的种子空种率也较高,平均50%左右。通过实际调查和资料查阅发现,血皮槭木质化的果皮坚硬而且很厚,不易透水、透气,种子深度休眠,发芽困难(Fordham,1969;杜丽雁等,2005),即使种子发育成幼苗,幼苗也比较脆弱。幼苗在所分布的群落中并非优势树种,林下光合有效辐射低,幼苗难以长大。
汪松等(2004)曾报导血皮槭种群数量正日趋减少,通过本文研究发现导致血皮槭种群数量减少的原因主要有以下几方面:野生血皮槭单株生长地地势险峻,生境恶劣,光照条件差,导致成熟单株结实量低或不结实;结实的种子空种率极高,从而导致土壤库中种子数量较少,种子受坚硬且很厚的果皮的限制自然条件下不易萌发,因此通过种子天然更新的幼苗便很少;天然更新幼苗的生境光照等条件不好,导致幼苗较难生长存活。本研究调查还发现,野生群体中血皮槭的大树数量也较少。除了上述自然因素,人为因素也是导致血皮槭野生资源数量逐年减少的另一原因。由于血皮槭材质坚硬树皮光滑观赏价值高,且由于树皮红色易发现,容易被砍伐盗用。上述综合因素导致血皮槭野生生境中的繁殖体数量较少,使其野生个体数量逐年减少。
| [1] |
陈丽,李冰,牛小沛,等.2012.血皮槭快繁过程中外植体褐化研究.现代农业科技,(8):198-198,206.( 1) 1)
|
| [2] |
陈丽,李冰,王梅,等.2010.血皮槭的愈伤组织培养研究.北方园艺,(8):170-172.(1)
|
| [3] |
杜丽雁,张春涛,高强,等.2005.血皮槭播种育苗技术.林业实用技术,(3):24.(2)
|
| [4] |
中国植物志编辑委员会.1981.中国植物志(第46卷).北京:科学出版社.(2)
|
| [5] |
付增娟,张川红,郑勇奇,等.2006.黑荆和银荆的繁殖扩散与入侵潜力.林业科学,42(10):48-53.(1)
|
| [6] |
李倩中,刘晓宏,苏家乐,等.2010.槭属种质遗传多样性及亲缘关系的SRAP分析.江苏农业学报,26(5):1032-1036.(1)
|
| [7] |
彭东辉,张启翔,郑世群,等.2006.福建茫荡山保护区野生蕨类资源及其园林应用.福建林学院学报,26(4):308-313.(1)
|
| [8] |
托比·马斯格雷夫,克里斯·加德纳,威尔·马斯格雷夫.2005.杨春丽,袁瑀译.植物猎人.广州:希望出版社.(1)
|
| [9] |
王沙生,高荣孚,吴贯明.1991.植物生理学.2版.北京:中国林业出版社.(1)
|
| [10] |
汪松,解焱.2004.中国物种红色目录.第1卷.红色目录.北京:高等教育出版社.(3)
|
| [11] |
许建伟,沈海龙,张秀亮,等.2010.花楸树种子散布、萌发与种群天然更新的关系.应用生态学报,21(10):2536-2544.(1)
|
| [12] |
张川红,郑勇奇,李继磊,等.2005.北京地区火炬树的萌蘖繁殖扩散研究.生态学报,25(5):978-985.(1)
|
| [13] |
张川红,郑健,郑勇奇.2008.特有植物血皮槭的研究进展∥郑勇奇.野生植物资源保护与可持续利用研究.北京:中国农业出版社,100-103.(1)
|
| [14] |
Halpern C B,Evans S A,Nielson S. 1999. Soil seed banks in young, close-canopy forests of the Olympic Peninsyla, Washington: potential contributions to understory reinitiation. Canadian Journal of Botany,77(7) : 922-955 .(1)
|
| [15] |
Moles A T,Drake D R. 1999. Potential contribution of the seed rain and seed bank to regeneration of native forest under plantation pine in New Zealand. New Zealand Journal of Botany,37:83-93 .(1)
|