 2012, Vol. 48
2012, Vol. 48文章信息
- 郭彦林, 孟庆繁, 高文韬
- Guo Yanlin, Meng Qingfan, Gao Wentao
- 长白山高山草甸植物-传粉昆虫相互作用网络可视化及格局分析
- Visulization and Pattern Analysis of Plant-Insect Pollinator Interaction Networks in Subalpine Meadow in Changbai Mountain
- 林业科学, 2012, 48(12): 141-147.
- Scientia Silvae Sinicae, 2012, 48(12): 141-147.
-
文章历史
- 收稿日期:2011-11-01
- 修回日期:2012-08-11
-
作者相关文章



2. 吉林省林业调查规划院 长春 130022
2. Jilin Institute of Forestry Inventory and Planning Changchun 130022
传粉昆虫在显花植物起源和演化的早期起着决定性的作用(任东等,1998),对现存有花植物的生存、繁衍也具有重要的作用。在显花植物中,大约有85%的物种属于虫媒传粉,利用传粉昆虫可显著地提高作物的产量。花通过提供花粉、花蜜等酬报以及休息、狩猎、觅偶交配和取暖的场所吸引访花昆虫访花,而昆虫在访花过程中或多或少地携带了花粉,触动了柱头,对所访的花产生授粉作用(钦俊德,1987)。
以往大多数植物与传粉昆虫物种间相互作用的研究仅关注种对种间或少数物种间高度特化的相互作用,如无花果与无花果小蜂间的相互作用,特定植物的传粉生态学等,很少涉及到群落水平上植物与传粉昆虫间的相互作用及群落组织结构的信息。事实上,群落中同时开花的植物与同时出现的传粉昆虫物种间存在着十分复杂的访问与被访问的关系网络,研究这些相互作用网络的结构特征,探索其形成的生态学过程及机制,有助于揭示物种间的协同进化关系(Bascompte et al., 2003;2006),为显花植物系统的保护提供理论支持。
近年来,网络理论和方法引入,为直观地描述和定量化分析复杂的植物-传粉昆虫互惠网络中物种间的相互作用提供了新工具(Bascompte et al., 2003;2006;Olesen et al., 2006;2008)。在网络理论中,把植物、传粉昆虫物种和2套物种间的相互作用抽象为图论中的顶(vertex)和边(edge)的关系,顶相当于计算机网络结构中的节点(node),边相当于计算机网络结构中的链接(link)。为此,可用包括2套顶和边的二部图(bipartite graph)来可视化地描述植物-传粉昆虫相互关系的拓扑结构(Jordano et al., 2003)。在拓扑结构中,套内的物种不发生联系,套间的物种相互联系(Pimm,2002)。二部图可以清楚地表明群落中哪些传粉昆虫访问了哪些植物物种,哪些植物受到了哪些昆虫的访问。
虽然植物、传粉昆虫相互关系二部图可以直观地描述植物和传粉昆虫物种间相互作用,为相互作用网络结构特征的研究分析提供便利,但当研究的植物-传粉昆虫相互作用网络所涉及到的物种过多、相互作用过于复杂时,难于在单一页面或单一屏幕上显示整个相互作用网络图,也难于查看和研究局部物种间的相互作用以及局部和局部间的相互关系。为此,需要引进一些新的方法。Pajek软件是由斯洛文尼亚卢布尔雅那大学开发的大型网络可视化和网络结构分析软件,可免费下载(http://vlado.fmf.uni-lj.si/pub/networks/pajek/) (de Nooy et al., 2005)。应用该软件提供的可视化平台多视角缩放功能在不同的水平和视角下构建直观的网络拓扑结构图,为在不同层面观察和分析网络组织结构提供了工具。
本文探索应用Pajek软件的多视角缩放功能分析不同分类阶元尺度和视角下的长白山亚高山草甸植物与传粉昆虫细化作用的网络结构,从不同层面揭示植物-传粉昆虫相互作用网络的组织机制。
1 研究地概况长白山自然保护区位于吉林省东南部,地理坐标为40°41′49″—42°25′18″N,127°42′55″—128°16′48″E,面积19.65万hm2。长白山自然保护区生态系统比较完整,自然环境复杂多样,动植物资源十分丰富,是我国最重要的温带森林生态系统保护区和生物多样性关键地区。1980年加入“人与生物圈计划(MAB)”,1986年被国务院批准为国家级自然保护区。
长白山亚高山草甸位于长白山自然保护区针叶林上缘和岳桦(Betula ermanii)林带(1 600~2 100 m),属于非地带性植被类型,显花植物种类丰富。大多数草甸植物在7—8月群花陆续相开放,丰富的显花植物和相对较长的开花时间支持了大量的传粉昆虫,为植物和传粉昆虫相互作用网络的研究提供了理想样本。
2 研究方法 2.1 调查时间野外调查工作于2008年进行,主要采用样带调查法。调查从6月中旬开始到10月上旬结束,每2周调查1次。调查时段为晴朗微风天气的上午9:00—12:00,下午2:00—5:00。
2.2 调查方法在长白山西坡,选择2块面积较大的有代表性的亚高山草甸作为调查样地,用GPS定位,在各样地内各选设20条1 m×20 m的平行样带。
显花植物调查:记录样带内所有显花植物的种类、花的大小、花的数量/单株植物、花型、花序结构、对称性、花冠颜色,各种植物(花)多度,花期。参照长白山植物图鉴、图谱现场鉴定显花植物种类,不能确定到种的采集标本、编号,带回实验室鉴定。
传粉昆虫调查:沿样带边线一侧记录样带内遇到的每株显花植物植株上访花昆虫的种类及其个体数量,拍照并摄像。野外能鉴定到种的昆虫,记录其种类,不能鉴定到种的,野外鉴定到属或科,按“形态种(morphospecies)”编号记录。记录每株植物上各访花昆虫种的个体数量(来访次数),并采集3~10头昆虫标本,分别按样地号、样带号、植物名称、采集日期、采集时段编号收集(Dicks et al., 2002;Gibson et al., 2006),用以存证标本、供实验室鉴定和虫体携带花粉检查。
2.3 分析方法1) 二部图构建长白山亚高山草甸植物-传粉昆虫拓扑关系图采用Pajek软件构建,其他图形采用SigmaPlot 10.0软件制作。
2) 物种度分析物种度(species degree)是指网络中某物种(顶点)与其他相关物种(顶点)发生联系的数量。在拓扑结构图中,从代表物种或高级分类阶元的顶中发出的连线的数量即代表了该物种或分类阶元的物种度的大小。物种度可以作为物种生态位宽度或特化水平的指标(Jordano et al., 2003;Bascompte et al., 2006;2007)。物种度的大小在很大程度上说明了该物种在整个网络结构中的重要性,物种度大的物种常常起到了网络中路由器(hub)的作用。从显花植物物种发出的连线越多,表明有越多的传粉昆虫物种来访,生态位越宽,反之越少,生态位越窄;同理,从传粉昆虫物种发出的连线越多,表明该种昆虫访问的植物物种越多。
为描述显花植物与传粉昆虫物种间相互作用网络, 定义矩阵
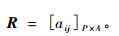
|
其中:P为显花植物物种数,A为传粉昆虫物种数;当植物物种i与传粉昆虫物种j间存在联系,则aij=1,否则aij=0。
根据相互关系矩阵R,定义
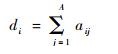
|
为第i种植物物种的物种度;同样,定义
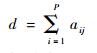
|
为第j种昆虫物种的物种度。
3 结果与分析 3.1 长白山亚高山草甸植物-传粉昆虫相互关系网络结构图的构建长白山亚高山草甸群落中,显花植物和传粉昆虫物种丰富,分类阶元组成多样,物种间访问关系复杂。要全面揭示这种复杂网络的组织机制和格局特征,不仅要直观地描述和考察群落中物种间的相互作用,而且要描述和考察目、科和属阶元尺度下的类群间的相互关系,甚至还要描述和考察大网络中局部类群物种间的相互作用。
根据网络结构理论,在不考虑各显花植物和传粉昆虫物种的种群密度及传粉昆虫的访问频度的情况下,物种和物种间的相互作用可以看成同等重要的顶和边。应用Pajek软件构建物种水平上的长白山亚高山草甸植物-传粉昆虫相互作用拓扑结构图,见图 1,应用软件提供的视角转换功能在图 1的基础上通过同一目的节点的归并构建显花植物-昆虫目相互作用拓扑结构图,见图 2;以及在局部视角下通过同一科的节点归并构建的植物-双翅目传粉昆虫相互作用拓扑结构图,见图 3。

|
图 1 长白山亚高山草甸显花植物-传粉昆虫相互作用拓扑结构 Fig.1 Topology of flowering plant-insect pollinator interaction network in subalpine meadow of Changbai Mountain |

|
图 2 显花植物-昆虫目相互作用拓扑结构 Fig.2 Topology of flowering plants-insects order interaction network |

|
图 3 显花植物-双翅目传粉昆虫相互作用拓扑结构 Fig.3 Topology of flowering plant-diptera interaction network |
在图 1~3中,左侧顶点代表植物物种,右侧顶点代表传粉昆虫物种或目,顶点之间的连线代表传粉昆虫物种或目与显花植物的访问关系。
3.2 植物-传粉昆虫相互关系网络结构格局分析物种度的频度分布是重要的网络格局特征之一,从物种度的频度分布分析可以洞察网络的组织机制。1) 物种水平上的植物-传粉昆虫相互作用网络结构格局物种水平上,长白山亚高山草甸中,植物-传粉昆虫相互作用网络由30种显花植物和101种传粉昆虫构成,物种间相互作用的拓扑结构见图 1。从图 4和图 5可见:显花植物和传粉昆虫的物种度频度分布基本上呈右偏态分布。

|
图 4 植物物种度频度分布 Fig.4 Frequency distribution of species degree for flowering plants in entire network |

|
图 5 昆虫物种度频度分布 Fig.5 Frequency distribution of species degree for insect pollinators in entire network |
显花植物物种度为1~80,平均为10.57±15.93。有1种以上昆虫访问的物种占76.67%,表明显花植物主要由生态泛化物种构成。物种度最大的是黑水当归(Angelica amurensis),物种度>20的显花植物物种还有长白老鹳草(Geranium baishanense)、续断菊(Sonchus asper)、返顾马先蒿(Pedicularis resupinata)和柳兰(Chamaenerion angustifolium)4种,占16.67%。物种度特高的超级泛化种缺失。
传粉昆虫物种度为1~17,平均为3.14±3.22。仅采访1种植物的传粉昆虫种占48.51%,与显花植物相比,传粉昆虫的生态特化大于显花植物物种。物种度超过10的昆虫物种仅有密林熊蜂(Bombus patagiatus)、散熊蜂(B. sprodicus)、黑带食蚜蝇(Episyrphus balteata)、黑珍蛱蝶(Clossiana hakulozana)、褐红珠灰蝶(Lycaeides subsolana)和长尾管蚜蝇(Eristalis tenax)6种,物种度>17的传粉昆虫物种也显著缺失。
2) 目水平上植物-昆虫目相互作用网络结构格局探索显花植物与传粉昆虫目间的相互作用对揭示传粉系统在更高分类阶元上的组织机制具有重要意义。
从图 6可以看出:在目的水平上,昆虫目的物种度频度分布不呈右偏态分布,与物种水平上物种度的频度分布格局不同。各目昆虫的物种度为1~26,平均13.38±7.85。各目昆虫物种度大小依次为膜翅目(Hymenoptera) (26)>双翅目(Diptera) (19)>鳞翅目(Lepidoptera) (18)>鞘翅目(Coleoptera) (16)>脉翅目(Neuroptera) (10)>直翅目(Orthoptera) (9)>半翅目(Hemiptera) (8)>蜻蜓目(Odonata)(1),在目等更高分类阶元尺度上显示了不同昆虫类群在传粉网络中的重要性。

|
图 6 昆虫目物种度的频度分布 Fig.6 Frequency distribution of species degree for insect order in plant-insect order network |
3) 植物-双翅目昆虫相互作用网络结构格局鉴于双翅目昆虫是长白山亚高山草甸中物种种类较多、多度较大的重要传粉昆虫类群,为此,从网络中提取显花植物-双翅目传粉昆虫相互作用的信息,探索在局部视角下植物与双翅目传粉昆虫物种间相互作用的拓扑结构格局。从图 7和图 8可以看出:在植物-双翅目传粉昆虫相互作用网络中,植物与双翅目传粉昆虫物种度的频度分布也呈右偏态分布。

|
图 7 植物-双翅目相互作用网络中昆虫物种度频度分布 Fig.7 Frequency distribution of species degree for insect pollinators in plant-Diptera network |

|
图 8 植物-双翅目相互作用网络中植物物种度频度分布 Fig.8 Frequency distribution of species degree for flowering plants in plant-Diptera network |
双翅目传粉昆虫物种度为1~12,平均3.07±3.15。物种度>1的传粉昆虫占51.85%。物种度超过10的昆虫物种仅有黑带食蚜蝇和长尾管蚜蝇2种,显花植物物种度为1~25,平均为4.37±5.45。受到1种以上昆虫访问的物种占68.42%。可见,从局部的视角看,显花植物与双翅目传粉昆虫也表现为主要由泛化物种构成,且物种度特高的超级泛化种显著缺失。
4 讨论利用Pajek软件提供的可视化平台构建了长白山植物-传粉昆虫相互作用网络整体拓扑结构图,并通过视角转换功能构建了不同分类阶元水平上的植物、传粉昆虫相互作用网络拓扑结构图,为在不同阶元水平上探讨尺度转换下植物-传粉昆虫相互作用网络组织机制的变化提供了途径。
从不同分类阶元水平上看,物种度的频度分布格局不同。在物种水平上,长白山亚高山草甸显花植物、传粉昆虫物种度的频度分布基本呈偏态分布,即网络中多数物种有少量的连接,少数物种有多数的连接,这与大多数传粉系统中物种度的分布一致(Waser et al., 1996;Jordano et al., 2003;Vázquez et al., 2003);但在目的水平上,传粉昆虫目的物种度频度分布并没有表现为上述格局。
物种度的频度分布显示了长白山亚高山草甸传粉系统中,显花植物、传粉昆虫物种生态位分化明显。表明在长期的协同进化过程中,一些植物倾向于通过生态泛化策略利用更多的传粉昆虫资源来增加传粉机会,而一些植物则倾向于利用专化性的传粉昆虫来提高传粉效率。传粉昆虫也通过2种策略更有效地获取酬报。
在物种水平上,利用1种以上的传粉昆虫或显花植物资源的物种所占的比例大,显示了大多数显花植物、传粉昆虫倾向于采取生态泛化的策略,为生态泛化种。群落中植物物种的平均物种度显著大于昆虫物种的平均物种度,这与传粉昆虫物种套中物种丰富度显著大于植物物种套中物种丰富度有关,与以往的研究结论一致(Waser et al., 1996)。物种度的频度分布格局也表明:无论传粉昆虫还是显花植物类群,均是由生态特化种到生态泛化种组成的连续统,且在连续统中缺乏超级泛化种,如在整个网络上,缺乏物种度大于80的显花植物物种及物种度大于18的传粉昆虫物种,这支持群落中存在生物物候和形态学限制的限制连接假说(forbidden links hypothesis)(Jordano et al., 2003;Rezende et al., 2007;Santamaría et al., 2007;Stang et al., 2007;Vázquez et al., 2009)。例如,传粉昆虫和花间物候期不重叠以及形态学不匹配阻止了2个物种发生相互作用。尽管在目的水平上,物种度的频度分布格局表现不同,但泛化的昆虫目占优势的特征与物种水平上的结论相一致。
物种度是物种在长期的进化过程中形成的物种特征,与物种生物地理学、形态学、生物气候学密切相关,也与局部种群密度和群落的组成相关。物种多度高的物种常常是花期长的植物和成虫活跃期较长的昆虫,是长白山亚高山草甸植物—传粉昆虫相互关系网络中的核心物种,是生物多样性保护的重点,也是生态系统退化后恢复的重点对象和以往的研究一样,本研究也是依据在有限的调查时间和有限的调查样地上取得的观察网络(observed network)上的调查数据,所得到的结论可能与真实的网络有一定的偏差(Vázquez et al., 2009;Olesen et al., 2011)。加大检查样地的数量和增加调查的时间,生态泛化种可能大幅增加,所得的结论更能接近真实状况。因此,如何减小抽样误差应该是未来研究需要关注的问题之一。
| [] | 钦俊德. 1987. 昆虫与植物的关系. 北京, 科学出版社. |
| [] | 任东, 洪友崇. 1998. 被子植物的起源——以喜花虻类化石为据. 动物分类学报, 23(2): 212–221. |
| [] | Barrat A, Lemy M B, Satorras R P, et al. 2004. The architecture of complex weighted networks. Proceedings of the National Academy of Sciences of the United States of America, 101(11): 3747–3752. DOI:10.1073/pnas.0400087101 |
| [] | Bascompte J, Jordano P, Melian C J, et al. 2003. The nested assembly of plant-animal mutualistic networks. Proceedings of the National Academy of Sciences of the United States of America, 100(16): 9383–9387. DOI:10.1073/pnas.1633576100 |
| [] | Bascompte J, Jordano P, Olesen J M. 2006. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science, 312: 431–433. DOI:10.1126/science.1123412 |
| [] | Bascompte J, Jordano P. 2007. Plant-animal mutualistic networks: the architecture of biodiversity. Annual Review of Ecology, Evolution, and Systematics, 38: 567–593. DOI:10.1146/annurev.ecolsys.38.091206.095818 |
| [] | Dicks L V, Corbet S A, Pywell R F. 2002. Compartmentalization in plant-insect flower visitor webs. Journal of Animal Ecology, 71(1): 32–43. DOI:10.1046/j.0021-8790.2001.00572.x |
| [] | de Nooy W, Mrvar A, Batagelj V.2005.Exploratory Social Network Analysis with Pajek.Structural Analysis in the Social Sciences 27. Cambridge University Press. |
| [] | Gibson R H, Nelson I L, Hopkins G W, et al. 2006. Pollinator webs, plant communities and the conservation of rare plants: arable weeds as a case study. Journal of Applied Ecology, 43: 246–257. DOI:10.1111/j.1365-2664.2006.01130.x |
| [] | Jordano P, Bascompte J, Olesen J M. 2003. Invariant properties in coevolutionary networks of plant-animal interactions. Ecology Letters, 6: 69–81. |
| [] | Olesen J M, Bascompte J, Dupont Y L, et al. 2006. The smallest of all worlds: pollination networks. Journal of Theoretical Biology, 240: 270–276. DOI:10.1016/j.jtbi.2005.09.014 |
| [] | Olesen E M, Bascompte J, Elberling H, et al. 2008. Temporal dynamics in a pollination network. Ecology, 89(6): 1573–1582. DOI:10.1890/07-0451.1 |
| [] | Olesen J M, Bascompte J, Yoko L, et al. 2011. Missing and forbidden links in mutualistic networks. Proccedings of the Royal Society of London, Series B, 278(1706): 725–732. DOI:10.1098/rspb.2010.1371 |
| [] | Pimm S L.2002.Food Webs. Second edition.The University of Chicago Press, Chicago, Illinois, USA. |
| [] | Rezende E, Jordano P, Bascompte J. 2007. Effects of phenotypic complementarity and phylogeny on the nested structure of mutualistic networks. Oikos, 116(10): 1919–1929. |
| [] | Santamara L, Rodrguez-Girones M. 2007. Linkage rules for plant-pollinator net-works: Trait complementarity or exploitation barriers?. PloS Biology, 5(2): 31. DOI:10.1371/journal.pbio.0050031 |
| [] | Stang M, Klinkhamer P G L, van der Meijden E. 2007. Asymmetric specialization and extinction risk in plant-flower visitor web: a matter of morphology or abunddence?. Oecologia, 151(3): 442–453. DOI:10.1007/s00442-006-0585-y |
| [] | Vázquez D P, Aizen M A. 2003. Null model analyses of specialization in plant-pollinator interactions. Ecology, 84: 2493–2501. DOI:10.1890/02-0587 |
| [] | Vázquez D P, Bluthgen N, Cagnolo L, et al. 2009. Uniting pattern an process in plant-animal mutulistic networks: a review. Annals of Botany, 103: 1445–1457. DOI:10.1093/aob/mcp057 |
| [] | Waser N M, Chittka L, Price M, et al. 1996. Generalization in pollination systems, and why it matter. Ecology, 77: 1043–1060. DOI:10.2307/2265575 |