 2012, Vol. 48
2012, Vol. 48文章信息
- 张彦龙, 杨忠岐, 王小艺, 吴成进, 马生福, 陆之贵
- Zhang Yanlong, Yang Zhongqi, Wang Xiaoyi, Wu Chengjin, Ma Shengfu, Lu Zhigui
- 2种昆虫病原线虫对松褐天牛幼虫的控制力测定
- Laboratory Test on Control of Japanese Pine Sawyer by Two Entomopathogenic Nematode Species
- 林业科学, 2012, 48(12): 64-70.
- Scientia Silvae Sinicae, 2012, 48(12): 64-70.
-
文章历史
- 收稿日期:2011-12-27
- 修回日期:2012-10-17
-
作者相关文章



2. 九华山风景区农村工作局 池州 242811
2. Rural Affair Bureau of Jiuhua Mountain Scenic Area Chizhou 242811
线虫属线形动物门(Nemathelminthes)线虫纲(Nematoda)。据统计,有1 000多种线虫寄生于700种以上的昆虫,仅被小卷蛾斯氏线虫(Steinernema carpocapsae)DD2136寄生的昆虫就有10目75科250种(Poinar,1979)。昆虫病原线虫作为一种生物防治因子已经应用于实践,如它们是许多土栖昆虫的重要天敌(Kaya et al., 1999),国际上较大面积应用昆虫病原线虫防治的害虫主要有根象甲类和草坪蝼蛄等(Parkman et al., 1992;Hudson et al., 1989;David et al., 2000),我国主要用于防治木蠹蛾类和出土期的桃小食心虫(Carposina niponensis)等隐蔽性害虫(杨怀文等,1999)。昆虫病原线虫中斯氏线虫(S. spp.)和异小杆线虫(Heterorhabditis spp.)拥有广泛的寄主,存在于农田、森林、草地、沙漠以及海滩等,对多种昆虫的寄生作用明显(Hominick et al., 1996)。
应用昆虫病原线虫控制天牛类蛀干害虫有过一些报道,其中应用最广的是斯氏线虫,对异小杆线虫的研究较少。在我国,卢希平等(1994)报道了异小杆线虫泰山Ⅰ号(H. sp.)、夜蛾斯氏线虫(S. feltiae)、斯氏线虫Agriotos品系对台湾柄天牛(Aphrodisium sauteri)、光肩星天牛(Anoplphora glabripennis)、桑天牛(Apriona germari)、黄带黑绒天牛(Embrikstrandia unifasciata)的幼虫的敏感性与寄生性。王希蒙等(1996)利用斯氏线虫的2个品系Agriotas和Beijing对光肩星天牛幼虫和蛹的侵染过程、天牛的死亡症状以及致死效应进行了研究。罗启浩等(1997)发现斯氏线虫A24能对钻蛀性害虫作寻向移动,日平均水平移动距离4.03 cm±0.72 cm。孙祥水(2000)利用斯氏线虫属的5个品系和异小杆线虫的1个品系对皱鞘双条杉天牛(Semanotus sinoauster)进行室内外试验,筛选出异小杆线虫泰山Ⅰ号为最佳品系。研究人员也将昆虫病原线虫用于松褐天牛(Monochamus alternatus)的防治。胡学难等(2006)在室内测定了夜蛾斯氏线虫对松褐天牛的致死中量和致死中时,同时将TDM模型引入试验结果分析。在日本,Mamiya(1989)利用不同浓度的2种斯氏线虫(S. kushidai和夜蛾斯氏线虫)对松褐天牛的幼虫和成虫进行感染,在培养基上和木段里面夜蛾斯氏线虫的杀天牛效果均明显好于S. kushidai,二者对成虫的杀死效果都在80%以上。Yamanaka(1994)利用斯氏线虫的Mexican品系侵染松褐天牛幼虫,6×106和12×106条·m-2剂量致死力分别为69.2%和72.2%,但是剂量远高于行业商品标准。Vega等(2000)对昆虫病原线虫的适宜温度进行了研究,15~25 ℃最合适。Phan(2008)还发现S. glaseri对松褐天牛毒性要好于小卷蛾斯氏线虫。目前国内外利用昆虫病原线虫防治天牛类害虫大多处在室内测试阶段,野外防治成功实例很少(潘洪玉等,2000),利用昆虫病原线虫防治松褐天牛的研究还处于起步阶段,需要测试出毒性较高的线虫品系,并进行林间试验。
松褐天牛是中国松树重要的蛀干害虫,能造成树势衰弱,甚至枯死,还是毁灭性森林病害——松材线虫(Bursaphelenchus xylophilus)病最主要的传播媒介(Morimoto et al., 1972;Mamiya et al., 1972;杨宝君等,1995)。松材线虫病对我国南方多省的松林危害严重,目前国内外还没有十分有效的方法控制此病。为探讨利用昆虫病原线虫防治松褐天牛进而控制松材线虫病的可行性,在室内测试了2种昆虫病原线虫对松褐天牛的控制能力。
1 材料与方法 1.1 材料线虫:小卷蛾斯氏线虫ALL品系,异小杆线虫(H. spp.)WG品系,购于广州市骏骊环保科技有限公司产品,固态培养基,规格为1亿条·(500 g)-1,保存在10 ℃的冰箱里待用。
松褐天牛:采自安徽九华山马尾松(Pinus massoniana)纯林中的诱木。挑选大小一致的个体,用解剖镜测微尺测量每头天牛前胸背板大小(前胸背板3.43~5.53 mm为3龄)(柳建定等,2008)。
木段:接天牛的木段采自九华山周围山上健康的马尾松,胸径0.12 m,截成长度0.35 m小段,两头用蜡封住,防止失水。
试验仪器:高0.40 m、口径0.20 m的单口玻璃筒,小型喷雾器,量筒、量杯等。
1.2 方法 1.2.1 试验处理试验前3天将天牛接入木段,用刀将木段的韧皮部撬开,再用凿子在已撬开的韧皮部下面木质部凿一个长10 mm、宽5 mm、深5 mm的小坑,将天牛放入小坑,再将韧皮部合拢、封严、编号。每个木段接30头3龄的松褐天牛幼虫。3天后,将2种线虫均配成4个浓度的悬浊液,浓度分别为3,4,5和6万条·mL-1。将配好的线虫悬浊液装入小型喷雾器内,直接喷洒在已接入天牛的木段表面,保证整个木段被药湿透。每个浓度处理3次重复, 共90头天牛。每天打开天牛的接入口,记录天牛生存状况,将死亡的天牛挑出来镜检是否由线虫致死,然后再封严接入口,连续观察7天。试验温度和相对湿度分别为25 ℃±2 ℃和75%±5%。
1.2.2 设置对照对照中所使用的木段天牛接入情况同1.2.1,放置在同样温湿度条件下,仅喷洒同量的蒸馏水。
1.2.3 数据处理所有数据均用DPSv6.50数据处理软件(唐启义等,2006),涉及到专业统计生物测定中的时间剂量死亡率模型。
1.3 利用TDM模型拟合时间-剂量-死亡率模型(time-dose-mortality model,TDM)分析方法正被用来替代Finney(1971)提出的概率分析法并逐渐为学术界所接受(Preisler et al., 1989)。TDM模型,按其数学结构又称互补重对数模型(complementary log-log model,CLL)。受试生物在时间tj(j=1,2,…,J)被供试因子在剂量di(i=1,2,…,I)下致死的概率可表达为
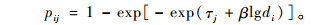
|
(1) |
式中,β被认为是描述“假设存在的每个关键生物受体所接受的活(毒)性分子的平均数目”,实际上可理解为描述剂量效应的斜率,τj是至时间tj的时间效应参数,τj+βlgdi部分称为线性预测因子(linear predictor)。式(1)中的pij是剂量di在第j个时间单位内产生的累计死亡概率(cumulative mortality probability)。由于累计死亡概率在时间上是连续的,若直接拟合则不符合模型模拟变量的独立性假设,因此转而考虑剂量di使受试个体在时间区间[tj-1,tj]内可能遭受的死亡率即条件死亡概率(conditional mortality probability),可表示为
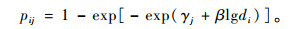
|
(2) |
式(2)β的含义与式(1)相同,γj为描述时间区间[tj-1,tj]内时间效应的待估参数,与τj在时间的含义上有所区别。由于条件死亡概率是独立的随机变量,所以式(2)才更适于进行模型的拟合。获得参数γj的估计后通过下式估计参数τj:

|
(3) |
γj和β的最大似然估计:通过二项分布最大似然函数

|
(4) |
的逼近,可获得γj和β的估计。式中,nij是在时间t1以剂量di处理ni头个体后在tj-1时刻仍然存活的个体数,rij是在时间区间[tj-1,tj]内死亡的个体数。但计算时,只需输入各剂量下每天的存活数和死亡数,即di,nij,rij(唐启义等,2006; 冯明光,1998)。
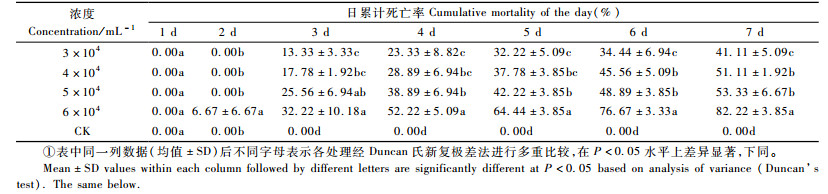
2 结果与分析 2.1 小卷蛾斯氏线虫ALL品系处理的天牛存活状况结果表明(表 1):ALL品系在3,4,5,6万条·mL-1的剂量下,经过7天松褐天牛3龄幼虫死亡率分别达41.11%,51.11%,53.33%,82.22%,对照没有出现死亡状况,因此死亡率无需校正。此线虫的持效能到第7天,6万条·mL-1的处理死亡率最高,达到82.22%。浓度为6万条·mL-1时,天牛从第2天就开始死亡,其余的浓度处理第3天才开始死亡。总体趋势是,随浓度增大累计死亡率也在升高;随时间的增加,累计死亡率增加;浓度最高的6万条·mL-1,开始死亡的时间最早,第2天死亡率6.67%。喷洒线虫第2天各个处理间松褐天牛累计死亡率间差异不显著(Df=4,14;F=3.001,P=0.07);第3天各个处理间松褐天牛累计死亡率间差异显著(Df=4,14;F=13.611,P=0.005);第4天差异显著(Df=4,14;F=28.305,P=0.000 1);第5天差异显著(Df=4,14;F=114.999,P=0.000 1);第6天差异显著(Df=4,14;F=115.501,P=0.000 1);第7天差异显著(Df=4,14;F=148.979,P=0.000 1)。
|
|
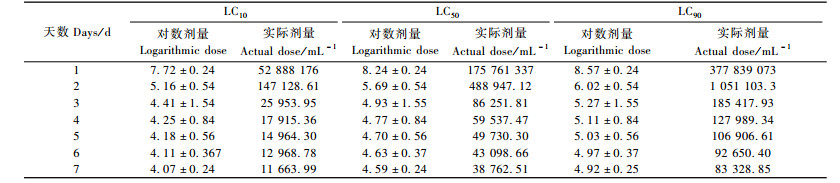
利用DPS软件处理表 1的数据,得出线虫ALL品系对松褐天牛3龄幼虫的TDM模型及对数剂量效应估计和对数剂量致死时间效应估计(表 2,3)。χ2=27.272 14(P=0.162 03) < χ2(20,0.05)=31.41,拟合模型达到显著水平。
|
|
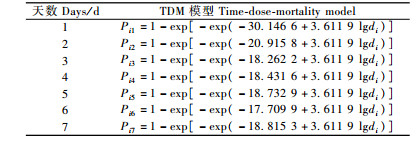
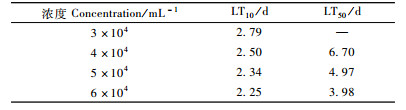
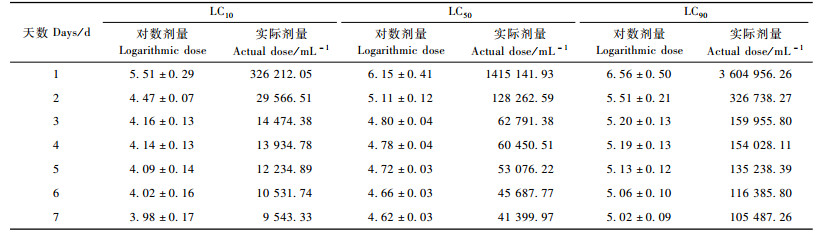
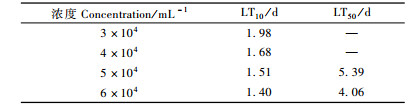
从表 2, 3可以看出:随着时间的延长,LC10, LC50和LC90逐渐减小,即时间越长,线虫与松褐天牛接触的机会就越多,使松褐天牛死亡一定数量所需的线虫剂量就越少;相反,时间越短就需要越大浓度的线虫。但是线虫制剂侵染和杀死天牛需要一定的时间,所以最初第1,2天不能仅仅用模型衡量,第1,2天的剂量(仅根据模型计算出来的浓度)是可能不切合实际的剂量。表 4中,随浓度的增大,致死一定数量的天牛所需的要的时间逐渐变短。3万条·mL-1的剂量的LT50没能由模型算出,是因为3万条·mL-1的处理样本的数据在最后一天的死亡率没能达到50%。
|
|
|
|
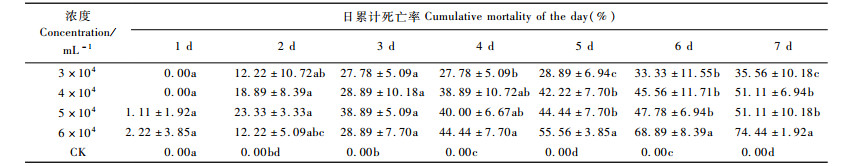
结果表明(表 5):WG在3,4,5,6万条·mL-1的剂量下,经过7天松褐天牛3龄幼虫死亡率分别达35.56%,51.11%,51.11%,74.44%,对照没有出现死亡状况,因此死亡率无需校正。此线虫的持效到第7天。6万条·mL-1的处理死亡率最高,达到74.44%。浓度为6万条·mL-1时,天牛从第1天就开始死亡,其余的浓度处理第2天才开始死亡。总体趋势是,随浓度增大累计死亡率也在升高;随时间的增加,累计死亡率增加;浓度为5,6万条·mL-1,开始死亡的时间最早,第2天死亡率分别为23.33%,12.22%。对不同浓度处理的日累计死亡率进行方差分析,结果显示喷洒线虫1天后各处理间松褐天牛累计死亡率差异不显著(Df=4,14;F=0.8,P=0.551 9), 2天后差异显著(Df=4,14;F=5.249,P=0.015), 3天后差异显著(Df=4,14;F=14.941,P=0.000 3),第4天差异显著(Df=4,14;F=19.827,P=0.000 1);第5天差异显著(Df=4,14;F=37.654,P=0.000 1);第6天差异显著(Df=4,14;F=24.742,P=0.000 1);第7天差异显著(Df=4,14;F=43.717,P=0.000 1)。
|
|
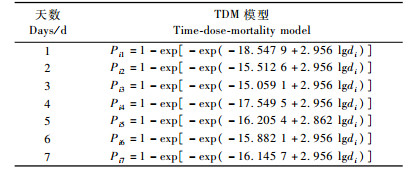
利用DPS软件处理表 1的数据,得出WG品系对松褐天牛3龄幼虫的TDM模型及对数剂量效应估计,对数剂量致死时间效应估计(表 6,7,8)。模型通过了Howierski等(1996)提出的Hosmer & Lemeshow卡方检验,χ2=13.338 1(P=0.100 73) < χ2(8,0.05)=15.51,模型达到显著水平。
|
|
|
|
|
|
从表 7可以看出:随着时间的延长,LC10, LC50和LC90逐渐减小,即时间越长,线虫与松褐天牛接触的机会就越多,使松褐天牛死亡一定数量所需的线虫剂量就越少;相反,时间越短就需要越大浓度的线虫。但是线虫制剂侵染和杀死天牛需要一定的时间,不仅仅符合模型就行,所以第1天的剂量(仅根据模型计算出来的浓度)是不切合实际的剂量。表 8中,随浓度的增大,致死一定数量的天牛所需要的时间逐渐变短。3万条·mL-1剂量的LT50没能由模型算出,是因为3万条·mL-1处理样本的数据在最后一天的死亡率没能达到或是刚到50%。
2.5 松褐天牛感染2种线虫日均死亡情况图 1, 2表明:ALL品系在第2天开始死亡,6万条·mL-1的死亡率为6.67%;第3天各个浓度处理均达到最高,4个浓度(3,4,5,6万条·mL-1)的死亡率分别为13.33%, 17.78%,25.56%,25.56%,然后随时间的延长,日均死亡率呈逐渐降低趋势。WG品系在第1天开始死亡,5,6万条·mL-1处理的日均死亡率分别为11.11%,22.22%;第2天4和5万条·mL-1达到最高, 分别是18.89%,22.22%;3,6万条·mL-1在第3天达到最高,分别为16.67%,15.56%,之后随时间的增长死亡率逐渐下降。二者的持续效果能到第7天,7天以后的数据未观测。显然WG品系的药效要比ALL品系快,二者的致死峰值主要集中在第1—3天。

|
图 1 松褐天牛幼虫被小卷蛾斯氏线虫All品系寄生后日均死亡率 Fig.1 The daily mean mortality of M. alternatus parasitized by S. carpocapsae ALL |

|
图 2 松褐天牛幼虫被异小杆线虫WG品系寄生后日均死亡率 Fig.2 The daily mean mortality of M. alternatus parasitized by H. sp. WG |
室内毒力测定表明ALL和WG品系线虫对松褐天牛有较强的致病性。
拟合的松褐天牛被2种不同品系的线虫侵染后的TDM模型以及估计的LC10,LC50,LC90,LT10,LT50等,对野外制剂选择、制剂浓度选择、防治时间选择等有指导作用。
2种线虫的起效时间不一样,WG品系在施药后24 h就开始致死天牛,而ALL品系在施药后48 h开始致死天牛;ALL品系起效慢但最终致死率高,比WG品系高7.78%,ALL品系药效强于WG品系。二者持续杀虫效果最低能达到7天。
3.2 讨论本次试验在研究方法上与以往的研究有所区别。首先,是选择天牛在进入木质部之前的龄期即3龄,此时天牛还处于低龄,体形较小,仅在木质部以外的韧皮部取食。Mamiya(1989)的研究证明用一些线虫防治已进入木质部的松褐天牛幼虫效果有限。其次,是利用木段接入法将3龄的天牛接入马尾松木段韧皮部位置,尽量模拟自然状态。以往的研究一般是直接利用培养皿或者是野外采回来的天牛已经进入木质部的木段,前者与野外条件差别太大,而后者选择的阶段不合适,这样很难检验出真实的效果。再次,利用喷洒树干的方法将线虫悬浊液施于木段上,这也是在野外最易实现的方法。以往的研究利用海绵塞孔法、侵入孔注射法、天牛浸泡法,这些方法在野外大规模实施存在一定局限。最后,生物测定以往一般使用传统的概率值分析(probit analysis)方法,即将累计死亡率进行几率值转换后分别对剂量或时间作线性回归分析,通过回归参数的估计而建立直线回归模型,从而估计剂量效应(LC50)或时间效应(LT50)。这样的分析方法使时间与剂量的效应相互分开,无法使所建模型充分体现试验数据的完整性。因此,有必要将时间和剂量的效应统一到同一个模型中来,本文利用TDM模型即将时间与剂量的效应进行了统一。
关于线虫致死天牛速度慢的可能原因。线虫被喷洒到木段上以后,需透过树皮缝隙寻找天牛,这会花费一定的时间;再通过天牛身上的自然孔进入天牛体内杀死天牛也需要时间。线虫在进入树皮过程中会遇见不可知的情况,如温度变低、大雨冲刷、其他生物捕食等,都有可能影响线虫的药效。
天牛所生存的环境湿度是个较大的限制因素。本文所说的湿度是指树皮含水量。线虫为湿生性生物,抗干燥能力弱,一般生活在湿度较大的土壤中,土壤水分不但影响线虫的生存,还影响线虫的活动(杨发柱,1989)。在向树上喷洒线虫制剂时,让树皮保持一定湿度,是保证防治效果的重要环节。雨后喷洒线虫制剂是否会提高防治效果,仍需深入研究。
光照也是防效的限制因素之一。当在松树的茎干上喷洒线虫后,强光的照射会对线虫产生影响,因此,应选择在下午或者阴天喷洒。光强度对线虫活动及生存情况的影响目前还没有研究,有待探索。利用线虫防治地下的害虫取得了一定成效,但是在田间大规模防治蛀干害虫,仍需要进一步研究。
其他生物对线虫的影响以及其对其他非靶标生物的影响不可忽视。1头真革螨(Eugomasus sp.)8 s能吞食1头DD-136线虫,1天能捕食110头;单齿线虫(Clarkus sp.)2 min吞食1头线虫,1天能捕食70头线虫;角咽线虫(Actinolaimus sp.)可以用口针刺吸DD-136线虫。自然界的情况复杂多变,其他生物对线虫以及线虫对其他生物防治的昆虫的影响有待于进一步研究。
2种线虫均可能对松褐天牛起到控制作用,需要浓度合适,条件适宜。要提高防治效果,就需要加大线虫剂量或延长时间。
| [] | 冯明光. 1998. 时间-剂量-死亡率模型取代几率分析技术. 昆虫知识, 35(4): 233–237. |
| [] | 郭郛, 忻介六. 1988. 昆虫学实验技术. 北京, 科学出版社: 377-382. |
| [] | 胡学难, 陈小帆, 阮乐秋, 等. 2006. 夜蛾斯氏线虫对松树上松褐天牛的毒力测定和室内防治效果. 中国生物防治, 22(1): 45–48. |
| [] | 柳建定, 李百万, 王菊英. 2008. 松褐天牛龄级划分方法初探. 浙江林业科技, 28(4): 84–86. |
| [] | 卢希平, 朱传祥, 刘玉. 1994. 利用病原线虫防治几种天牛幼虫的室内寄生性测试. 森林病虫通讯(4): 31–33. |
| [] | 罗启浩, 谭长青, 陈志凌, 等. 1997. 昆虫病原线虫防治木蠹蛾和天牛幼虫的研究. 华南农业大学学报, 18(1): 25–30. |
| [] | 潘洪玉, 席景会, 张浩. 2000. 昆虫病原线虫对隐蔽性害虫控制作用的研究. 吉林农业大学学报, 22(1): 41–44. |
| [] | 孙祥水. 2000. 利用昆虫病原线虫防治皱鞘双条杉天牛. 福建林学院学报, 20(1): 49–51. |
| [] | 唐启义, 冯明光. 2006. DPS数据处理系统--实验设计、统计分析及模型优化. 北京, 科学出版社: 295-304. |
| [] | 王希蒙, 马峰, 任国栋. 1996. 昆虫病原线虫对天牛致死效应的室内试验. 宁夏农学院学报, 17(3): 5–8. |
| [] | 杨宝君, 朱克恭, 周元生. 1995. 中国松材线虫病的的流行与治理. 北京, 中国林业出版社: 113-114. |
| [] | 杨发柱. 1989. 昆虫病原线虫应用的生态学问题. 昆虫天敌, 11(3): 142–145. |
| [] | 杨怀文, 陈松笔. 1999. 昆虫寄生线虫与林业害虫的防治. 林业科学, 35(6): 103–110. |
| [] | David I S, Clay W M. 2000. Virulence of entomopathogenic nematodes to Diaprepes abbreviatus (Coleoptera: Curculionidae) in the laboratory. J Econ Entomol, 93(4): 1090–1095. DOI:10.1603/0022-0493-93.4.1090 |
| [] | Finney D J. 1971. Probit Analysis. 3rd ed. Britain, Cambridge University Press: 70-83. |
| [] | Hominick W M, Reid A P, Bohan D A, et al. 1996. Entomopathogenic nematodes: biodiversity, geographical distribution and the convention on biological diversity. Biocontrol Sci Technol, 6: 317–331. DOI:10.1080/09583159631307 |
| [] | Hudson W G, Nguyen K B. 1989. Infection of Scapteriscus vicinus (Orthoptera: Gryllotalpidae) nymphs by Neoaplectana sp. (Rhabditida: Steinernematidae). Florida Entomologist, 72: 383–384. DOI:10.2307/3494924 |
| [] | Kaya H K, Koppenhöfer A M. 1999. Biology and ecology of insecticidal nematode//Polavarapu S. Optimal Use of Insecticidal Nematodes in Pest Management. New Jersey: Rutgers University Press, 1-8. |
| [] | Mamiya Y. 1989. Comparison of the infectivity of Steinernema kushidai (Nematode: Steinernematidae) and other steinernematid and heterorhabditid nematodes for three different insects. Appl Ent Zool, 24(3): 302–308. DOI:10.1303/aez.24.302 |
| [] | Mamiya Y, Enda N. 1972. Transmission of Bursaphelenchus lignicolus (Nematode: Aphelenchoidae) by Monochamus alternatus (Coleoptera: Cerambycidae). Nematologica, 18(4): 159–162. |
| [] | Morimoto K, Iwasaki A. 1972. Role of Monochamus alternatus (Coleoptera: Cerambycidae) as a vector of Bursaphelenchus lignicolus (Nematoda: Aphelenchoididae). Journal of Japanese Forest Society, 54(6): 177–183. |
| [] | Nowierski R M, Zeng Z, Stenfan J, et al. 1996. Analysis and modeling of time-dose-mortality of Melanoplus sanguinipes, Loscusta migratoria migratorioides and Schistocerca gregaria (Orthoptera: Arididae) from Beauveria, Metarhizium and Paecilomyces isolates from Madagascar. J Invertebr Pathol, 67(3): 236–252. DOI:10.1006/jipa.1996.0039 |
| [] | Parkman J P, Frank J H. 1992. Infection of sound-trapped mole crickets, Scapteriscus spp., by Steinernem scapterisci. Florida Entomologist, 75(1): 163–165. DOI:10.2307/3495500 |
| [] | Phan Long Ke. 2008. Potential of entomopathogenic nematodes for controlling the Japanese pine sawyer, Monochamus alternatus//Zhao B G, Futai K, Sutherland J R, et al. Pine Wilt Disease. Japan: Springer, 371-381. |
| [] | Poinar G O. 1979. Nematodes for biological control of insect. Florida, CRC Press: 2-11. |
| [] | Preisler H K, Robertson J L. 1989. Analysis of time-dose-mortality data. J Econ Entomol, 82(6): 1534–1542. DOI:10.1093/jee/82.6.1534 |
| [] | Vega F E, Lacey L A, Reid A P, et al. 2000. Infectivity of a bulgarian and an American strain of Steinernema carpocapsae against codling moth. Biocontol, 45: 337–343. DOI:10.1023/A:1009979222466 |
| [] | Yamannka S. 1994. Field control of Japanese pine sawyer, Monochamus alternatus larvae by Steinernema carpocapsae. Jpn J Nematol, 23(2): 71–78. |