 2012, Vol. 48
2012, Vol. 48文章信息
- 尚莉莉, 孙正军, 江泽慧, 刘杏娥, 杨淑敏
- Shang Lili, Sun Zhengjun, Jiang Zehui, Liu Xing'e, Yang Shumin
- 毛竹维管束的截面形态及变异规律
- Variation and Morphology of Vascular Bundle in Moso Bamboo
- 林业科学, 2012, 48(12): 16-21.
- Scientia Silvae Sinicae, 2012, 48(12): 16-21.
-
文章历史
- 收稿日期:2012-09-18
- 修回日期:2012-10-24
-
作者相关文章



竹材是可迅速再生的生物材料之一,力学性能好,强度和模量超过木材;断裂韧性和疲劳性能超过多数的工程材料(Jiang, 2002; Amada et al., 2001; Nogata et al., 1995; Ding et al., 1995)。竹材主要由维管束和基本组织构成,维管束是主要的承载部分。维管束由外纤维鞘、内纤维鞘、侧生纤维鞘、初生韧皮部、初生木质部等组织构成。维管束是竹秆的主要承重单元,其特征对竹子的分类有着重要意义。20世纪30年代,日本学者竹内叔雄等对竹材维管束的形态进行了初步的研究(林新春等,2005),随后,国内外学者对竹材维管束的结构展开了系统全面的研究,并分别提出了相关的分类方法(李正理等,1960;1962;腰希申等,1993;朱慧芳等,1964; Grosser et al., 1971)。然而,对于竹材维管束截面面积的精细研究却鲜有报道。多数学者在研究竹材维管束性质时,仅利用普通光学显微镜对维管束面积进行粗略测量。例如:Amada等(1996)用CCD摄像机和图像分析软件测量了毛竹(Phyllostachys edulis)纤维的体积分数;Li等(2011)用Leica DM2500光学显微镜测量了毛竹维管束的面积,面积值文中未给出;王军等(2009)利用OLYMPUS BX41偏光显微镜和OLYMPUS C5050数码相机获取了毛竹秆茎维管束横切面结晶图像。这些测量方法虽然比较简便,但由于测试手段的局限,难以有效而准确地测量维管束的面积。本文利用激光共聚焦显微镜精确测定了毛竹维管束及其各组成单元的面积,提出了一种测量竹材维管束面积的新方法。此种方法相比前人的测试方法主要有2个优点:一是用于测量的切片厚不足2 μm,可保证维管束的立体形状;二是在激光共聚焦下能清楚地观察到纤维和薄壁细胞的界面,保证了测量的精确性。本文还研究了内、外层维管束的面积的变化规律,为后续研究维管束力学性能奠定基础,使将竹材的微观力学性能和宏观力学性能联系起来成为可能。
1 材料与方法 1.1 试验材料毛竹,4年生,采自安徽省黄山公益林场,胸径99 mm,秆长7 760 mm,共30节,分为3段,详细情况见表 1。
|
|
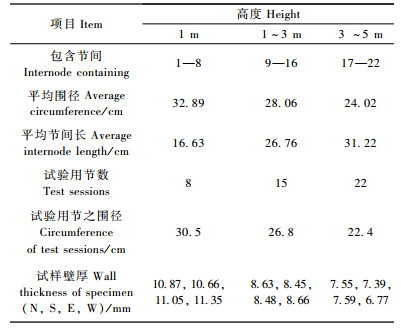
将试验用的竹筒(长约220 mm)分别沿南、北向劈成宽20 mm左右的竹条;从竹条上截取100 mm长的南北向竹段各1个;用刀将100 mm长竹段的竹壁按1/3竹壁厚度分割成内(竹黄)、中(竹肉)、外(竹青)3部分薄片。
薄片经水煮软化后剥取维管束,内部剥取10根,中部和外部剥取15根。从剥取的维管束上截取2 cm长的一段作为面积测量样品。维管束先用酒精脱水,然后在包埋管内滴入适量的Spurr树脂,将维管束插入其中,并保持其竖直。树脂只会填充到导管和细胞腔中起支撑作用,不会改变细胞壁的结构(Spurr, 1969)。
1.2.2 面积测量将维管束样品用超薄切片机切取1.5 μm厚的切片,每个维管束样品切出的薄片数不少于10片。在光学显微镜下挑出质量最好的3片,经吖啶橙染色后,封存在载玻片上。用激光扫描共聚焦显微镜(LSM 510 Meta,Zeiss)扫描,获取维管束横截面图片并测量面积。
1.3 维管束各组分定义将竹材沿竹秆纵向定义为竹长(L),沿圆周方向定义为弦向(T),沿竹壁厚度方向定义为径向(R)(Ghavami et al., 2003)。竹材的各方向定义见图 1。

|
图 1 竹材各方向的定义(N北向;S南向;T弦向;L纵向;R径向) Fig.1 Assumed axis in respect to longitudinal (L), tangential (T), and radial (R) directions, north(N)and south(S) |
将维管束沿内外纤维鞘方向定义为径向(R),又称长轴;沿侧方纤维鞘方向定义为弦向(T),又称短轴。毛竹维管束的方向定义见图 2。

|
图 2 维管束的方向及各部分定义 Fig.2 Assumed vascular bundle in respect to tangential (T) and radial (R) directions, and defined the fibrous sheath(f), porous(p), vessel(v) |
毛竹秆横切面维管束的分布见图 3。一般是位于外侧的小而密,位于内侧的大而疏。典型维管束通常位于竹秆横切面的中部或内部,按斜行排列并具有多行。

|
图 3 毛竹竹壁横切面宏观结构 Fig.3 The macroscopic structure in cross section of Moso bamboo wall |
在同一切面,竹材外层、中层和内层的维管束形态和大小变化明显。竹壁横切面的不同部位的维管束在分布上有显著差异。在竹壁外层的维管束分布密集,但尺寸较小,输导组织分化不完全,纤维细胞是主要成分。横切面中部的维管束形态稳定(变化不大),且输导组织分化完全,是竹种的特征形状(葛罗逊等, 1982)。毛竹维管束形态变化模式见图 4。

|
图 4 毛竹维管束从外到内的变化(4年生,3 m处) Fig.4 The deformation of Moso bamboo vascular bundles from outer to inner (4-year-old, at 3 m) |
从图中可以看出,毛竹横截面维管束存在2种形态,一种为半开放型维管束,另一种为开放型维管束。由内到外按2种形态可将竹壁分为内、外2层,竹青属于外层,竹黄属于内层,竹肉一部分属于内层,一部分属于外层,且在竹秆不同部位分属内、外层的比例不同,具体的比例还需要试验进一步摸索。半开放型维管束分布于横切面的外层,开放型维管束分布于横切面的内层。在外层,维管束排列有明显的规律,呈错列排列,形态由小逐渐变大;在内层,维管束形态一致,但靠近外层的维管束长宽比大;从外向里,维管束的长轴逐渐变小,短轴变大,长宽比逐渐减小;靠近髓腔,长轴和短轴几乎相等。
重新分类后的内层维管束和外层维管束形态见图 5。从图 5中可以看出,内层维管束包含2种形态:一种是4个纤维鞘相互独立,如图 5A所示;另一种是维管束的下方纤维鞘与其中的一个侧方纤维鞘相连,而另一个侧方纤维鞘和外方纤维鞘相互独立,即维管束包含3个独立的纤维鞘,如图 5B所示。外层维管束在形态上比较一致,都只有2个独立的纤维鞘,但纤维鞘大小有差别,如图 5C, D所示。

|
图 5 内层和外层维管束的形态 Fig.5 The morphology of vascular bundles in the inner layer and outer layer A, B.内层维管束形态Inner vascular bundles morphology; C, D.外层维管束形态Outer vascular bundles morphology. |
根据维管束的组成,将导管、初生韧皮部和薄壁细胞统称为综合多孔相,将纤维鞘看成是纤维相,故而可以把维管束看成是纤维相和综合多孔相的复合体。因此维管束的面积测量包括维管束的外围面积的测量、导管孔的面积测量、综合多孔相和纤维相的面积测量。
将维管束的面积定义为Av,综合多孔相的面积定义为Ap,纤维相的面积定义为Af。根据复合材料定律,则有下式成立:Av=Af+Ap, 即纤维相的面积可由维管束面积和综合多孔相的面积计算得到。典型维管束的面积测量见图 6。

|
图 6 毛竹维管束的面积测量
Fig.6 The area measurement of Moso bamboo vascular bundles
A.内层Inner layer;B.外层Outer layer. A:总面积Area of vascular; Av:导管面积Area of vessel; Af:竹纤维鞘面积Area of fibrous sheath; Ap:多孔相面积Area of porous. |
毛竹维管束的面积为0.038 3~0.246 2 mm2,平均值为0.155 5 mm2;导管孔的面积为0~0.058 5 mm2,平均值为0.022 0 mm2;综合多孔相的面积为0.000 5~0.147 8 mm2,平均值为0.055 9 mm2;纤维鞘的面积为0.037 8~0.143 6 mm2,平均值为0.099 6 mm2(图 7)。

|
图 7 毛竹维管束面积值 Fig.7 The statistics of area of the Moso bamboo vascular bundles |
维管束的形态在同一秆茎的不同高度(轴向)及同一高度的不同部位(径向)不同,因而其面积也有差异(图 8)。

|
图 8 毛竹维管束及其组成单元的面积变异规律 Fig.8 Variation of area of the Moso bamboo vascular bundles and component units |
从图中可以看出,毛竹维管束的横截面面积沿轴向变化不明显,但在径向变化明显,与维管束的大小变化一致。前人的研究也表明,不同高度的维管束大小差别不大。外层维管束的各组分面积和内层维管束的各组分面积相比都要小很多,特别是导管面积和多孔相面积。内层维管束的导管面积是外层维管束导管面积的2.59倍,其多孔相面积是外层维管束多孔相面积的4.15倍,这一点也证明了在形态上,外层维管束中输导组织未分化完全。虽然外层维管束的面积小于内层维管束,但外层维管束纤维鞘面积与内层维管束纤维鞘面积相差不大,故外层纤维体积分数大于内层,进一步证实了在竹壁横切面上维管束的体积分数从内向外增加(Amada et al., 1996)。而纤维是竹材中主要的承载结构,研究证明竹子纤维的拉伸强度约为基体的10倍(Amada et al., 1997),这从另外一面证明了竹材中力学强度是竹壁的外侧大于内侧。竹材的这一结构是为了抵抗自然界中频繁的风载。
2.3 毛竹维管束的面积变化规律维管束在内层和外层的排列分布不同,其面积规律也不同。外层维管束排列紧密,在横切面上呈错列排列,形态较小,面积值也有规律的变化。
在外层第1排,维管束面积小,无导管和薄壁细胞等其他组织;第2排开始形成完整的维管束,其径向长度增加,面积增大;第1, 2排通常是未分化的维管束,其输导组织的面积极小,厚壁纤维细胞构成的束状组织排列十分紧密,形成竹秆坚韧的外壁。从第2排到第9排,外层维管束的面积规律见图 9,面积值随径向位置的变化总体呈增加趋势。分析发现,纤维鞘面积随径向位置增加是维管束面积增加的主要因素,且纤维鞘面积与径向位置(从外到内)有显著的正相关线性关系,前人的研究(Li et al., 2011)证明纤维体积分数随着径向位置从内向外变化而增加。多孔区面积和导管面积也与径向位置有很好的正相关线性关系。故整体表现上维管束的面积随径向位置从内向外变化而增大。

|
图 9 内、外层维管束的面积变异规律(4年生,3米处) Fig.9 Area variation of outer and inner vascular bundles (4-year-old, at 3 m) |
内层维管束在横切面上分布比较均匀,维管束形态较大,其面积差异小。就各组分面积来说,导管孔面积和多孔区面积大,纤维鞘面积占总面积的比例小。分析发现,导管面积与径向位置有正相关线性关系,而纤维面积随径向位置增加而略有减小。所以整体表现维管束面积差异不大。
3 结论竹壁由内到外按维管束形态可划分为内层和外层2层。毛竹维管束的面积为0.038 3 ~0.246 2 mm2;导管孔的面积为0~0.058 5 mm2;综合多孔相的面积为0.000 5~0.147 8 mm2;纤维鞘的面积为0.037 8~0.143 6 mm2。毛竹外层维管束排列紧密,在横切面上呈错列排列,维管束形态较小,面积值随径向位置的变化总体呈增加趋势;内层维管束在横切面上分布比较均匀,维管束形态较大,其面积差异小。就各组分面积来说,导管孔面积和多孔区面积大,纤维鞘面积占总面积的比例小。
| [] | 葛罗逊, 里斯. 1982. 关于亚洲竹类的解剖及其维管束. 耿伯介, 译. 竹子研究汇刊, 1(1): 105-130. |
| [] | 林新春, 方伟. 2005. 竹亚科分类学研究进展. 竹子研究汇刊, 24(1): 1–6. |
| [] | 李正理, 靳紫宸, 腰希申. 1960. 几种国产竹材的比较解剖观察. 植物学报, 8(1): 25–30. |
| [] | 李正理, 靳紫宸, 腰希申. 1962. 几种国产竹材的比较解剖观察续报. 植物学报, 10(1): 15–38. |
| [] | 王军, 王伟军, 刘云飞, 等. 2009. 毛竹秆茎维管束结晶图像的分割. 东北林业大学学报, 37(1): 105–106. |
| [] | 腰希申, 梁景森, 马乃训, 等. 1993. 中国主要竹材微观构造. 大连, 大连出版社. |
| [] | 朱慧芳, 腰希申. 1964. 国产33种竹材制浆应用上纤维形态结构的研究. 林业科学, 9(4): 311–331. |
| [] | Amada S, Munekata T, Nagase Y, et al. 1996. The mechanical structures of bamboos in viewpoint of functionally gradient and composite materials. Journal of Composite Materials, 30(7): 800–819. DOI:10.1177/002199839603000703 |
| [] | Amada S, Ichikawa Y, Munekata T, et al. 1997. Fiber texture and mechanical graded structure of bamboo. Composites Part B: Engineering, 28(1/2): 13–20. |
| [] | AmadaS, Untao S. 2001. Fracture properties of bamboo. Composites Part B: Engineering, 32(5): 451–459. DOI:10.1016/S1359-8368(01)00022-1 |
| [] | Ding Y, Liese W. 1995. On the nodal structure of bamboo. Journal of Bamboo Research, 14(1): 24–32. |
| [] | Ghavami K, Allameh S M, Sánchez M L, et al. 2003. Multiscale study of bamboo Phyllostachys edulis. João Pessoa, Brazil: Proceeding of IACNOCMAT. |
| [] | Grosser D, Liese W. 1971. On the anatomy of Asian bamboos, with special reference to their vascular bundles. Wood Science and Technology, 5(4): 290–312. DOI:10.1007/BF00365061 |
| [] | Jiang Zehui. 2002. Bamboo and Rattan in the World. Shenyang, China, Liaoning Science and Technology Publishing House. |
| [] | Li Hongbo, Shen Shengping. 2011. The mechanical properties of bamboo and vascular bundles. Materials Research Society, 26(21): 2749–2756. DOI:10.1557/jmr.2011.314 |
| [] | Nogata F, Takahashi H. 1995. Intelligent functionally graded material: Bamboo. Composites Engineering, 5(7): 743–751. DOI:10.1016/0961-9526(95)00037-N |
| [] | Spurr A R. 1969. A low-viscosity epoxy resin embedding medium for electron microscopy. Journal of Ultrastructure Research, 26(1/2): 31–43. |