 2012, Vol. 48
2012, Vol. 48文章信息
- 李伟, 崔丽娟, 张守攻
- Li Wei, Cui Lijuan, Zhang Shougong
- 太湖岸带湿地不同植被覆被条件下土壤种子库的时空异质性
- Temporal and Spatial Heterogeneity of Soil Seed Bank Under Different Vegetation Cover Types in Lakeshore Wetland, Lake Tai
- 林业科学, 2012, 48(12): 10-15.
- Scientia Silvae Sinicae, 2012, 48(12): 10-15.
-
文章历史
- 收稿日期:2011-08-11
- 修回日期:2012-09-04
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091
2. Research Institute of Forestry, Chinese Academy of Forestry Beijing 100091
土壤种子库(soil seed bank)指存在土壤表面和土壤中所有具有活力的种子总和(Thompson et al., 1979)。种子通过种子雨进入土壤种子库,物种成熟节律和种子散落规律首先影响着土壤种子库的季节变化,然后随着种子的萌发、死亡或被摄食而形成一个动态的过程(唐樱殷等, 2011),种子库是决定植物群落物种多样性及其动态的重要因素(李伟成等, 2007; 葛斌杰等, 2010)。作为植物群落的潜在物种库,种子库对于种群个体的繁殖、扩散、恢复以及对不良环境的抵抗均有着重要意义(Rossell et al., 1999; 沈泽昊等, 200; 尹华军等, 2005),也是植物群落物种多样性及其动态的重要决定因素(Bossuyt et al., 2009)。土壤种子库的空间变化可以对地表植物群落的形成产生影响(Tekle et al., 2000; Vecrin et al., 2007)。目前,国内陆续开展了一些湿地土壤种子库的研究,例如:长江中下游湿地(陈中义等, 2001; 刘贵华等, 2007;叶春等, 2008; 侯志勇等, 2009; 徐洋等, 2009; 李守淳等, 2011; Lu et al., 2012),三江平原湿地(王正文等, 2002; 邢福等, 2008)和塔里木河流域湿地(徐海量等, 2007; 李吉玫等, 2008)等。土壤种子库研究在岸带湿地以及湖滨带开展得较少(徐洋等, 2009; 李守淳等, 2011),主要针对土壤种子库的特征,而太湖岸带湿地种子库时间、空间分布特征鲜有报道。
岸带湿地具有维持生物多样性、净化污染、减少洪涝灾害等功能,能够为生物物种提供良好的栖息环境,通过过滤、渗透、吸收、滞留沉积物质和能量,减缓径流,减弱进入地表和地下水的污染物毒性,降低污染程度(王超等, 2003; 夏继红等, 2006; Wang et al., 2007)。长期以来太湖岸带作为防洪大坝而不断加高加固,太湖岸带湿地受人为干扰严重,垦殖现象严重,河网水系复杂,岸带土壤受到的影响较为明显。本文以太湖岸带湿地为研究对象,系统分析了不同覆被类型土壤种子库的物种组成、分布格局及季节变化,旨在探讨岸带湿地种子库的特征及存在机制,为太湖岸带湿地植被恢复提供理论基础。
1 研究区概况太湖位于长江三角洲南缘,分属江苏、浙江、上海两省一市,位于30°56′—31°34′ N,119°54′—120°36′ E。南北最大长度为68 km,东西最大宽度为56 km,太湖全水域面积2 425 km2,为我国第3大淡水湖,是江苏最大湖泊(Wang et al., 2007)。属北亚热带湿润性季风气候,受太湖水体调节作用影响,四季分明,温暖湿润,雨水丰沛,光照充足,全年平均气温为17 ℃,年平均降雨量1 096.9 mm。主要气候特征如下:极端最低气温-6.6 ℃;极端最高气温36.8 ℃;年平均降水日数133.9天;年平均日照时数2 005~2 179 h;年平均相对湿度79%;常年主导风向为东南风,次主导风向为西北风(Hu et al., 2011)。在研究期内,太湖水位无明显的季节变化,植物生长季节其水位相对稳定。
研究区集中于太湖东岸,平均海拔4 m左右,地上植被优势种包括芦苇(Phragmites australis)、芦竹(Arundo donax)、香蒲(Typha orientalis)、喜旱莲子草(Alternanthera philoxeroides)、水蓼(Polygonum hydropiper)、葎草(Humulus scandens)和酸模(Rumex acetosa)等。其中,酸模岸带靠近太湖湿地公园,主要由酸模、葎草、稗(Echinochloa crusgalli)、飞蓬(Erigeron acer)、狗尾草(Setaria viridis)、野大豆(Glycine soja)、芦苇、藜(Chenopodium album)、大车前(Plantago major)和酸模叶蓼(Polygonum lapathifolium)等物种组成;灌草岸带主要由柽柳(Tamarix chinensis)、垂柳(Salix babylonica)、喜旱莲子草、芦苇、水蓼、菰(Zizania latifolia)、香蒲、假稻(Leersia japonica)、苦荬菜(Ixeris polycephala)、铁苋菜(Acalypha australis)、酸模叶蓼、稗和菟丝子(Cuscuta chinensis)等物种组成;农作物岸带主要是当地居民在岸带区开辟种植的农作物,主要包括番薯(Ipomoea batatas)、芝麻(Sesamum indicum)、豆类等作物,伴生植物包括葎草和灰绿藜(Chenopodium glaucum)等;天然芦苇岸带主要以芦苇和喜旱莲子草为优势种,伴生物种包括芦竹、野大豆、荠(Capsella bursapastoris)、葎草、水蓼和萝藦(Metaplexis japonica)等。
2 研究方法 2.1 种子库取样与萌发2010年4,8和11月,在试验区内随机选取酸模岸带(A区)、灌草岸带(B区)、农作物岸带(C区)、天然芦苇岸带(D区)斑块各5个。每个岸带斑块内用取土器取5个土样,取样面积为10 cm×5 cm,取样深度15 cm,分3层(0~5 cm,5~10 cm,10~15 cm),装袋带回实验室。
用幼苗萌发法估计种子库的种子数量。样品带回实验室后,将取自同一斑块的5个土样混匀,然后平铺到10 cm×5 cm×5 cm的萌发盆中,重复5次。为使种子库中的种子充分萌发,萌发过程中,所有萌发样品光照充足,每天适时补充水分以保持人工气候箱内湿润。白天温度30 ℃,时间14 h;晚上温度25 ℃,时间10 h,空气湿度保持在60%。每周记录1次萌发的物种种类及数量,幼苗一经鉴定即移走,暂不能鉴定的幼苗保留至可鉴定为止。
2.2 数据处理将取样面积内的种子数量换算成1 m2的数量,以lg(x+1)进行对数转换后,满足方差齐性。利用Spss18.0软件运用单因素方差分析(One way ANOVA)检测不同覆被类型及不同层次种子库密度的差异。
相似性采用Sorensen相似性系数(similarity coefficient,SC)计算:
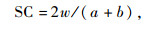
|
式中:w为共有物种数;a,b为某种类型种子库中的物种数。
3 结果与分析 3.1 土壤种子库物种组成由表 1可知,在A区酸模岸带、B区灌草岸带、C区农作物岸带、D区天然芦苇岸带4种植被覆被下的3期试验中共计检出30个物种。2010年4,8和11月分别萌发21,16和18个物种。从不同生活型物种组成来看,1年生和多年生草本占所有萌发物种的90%以上。其中,稗、碎米荠(Cardamine hirsuta)、酸模、喜旱莲子草,天胡荽(Hydrocotyle sibthorpioides)、芦苇、水莎草(Juncellus serotinus)、鼠麹草(Gnaphalium affine)、湿地勿忘草(Myosotis caespitosa)、牛筋草(Eleusine indica)、蔊菜(Rorippa indica)等11种植物在3个调查时段内都有出现。针对不同覆被类型来看,在A区岸带,物种酸模在3个时段皆有出现;B区岸带喜旱莲子草、鼠麹草、湿地勿忘草、天胡荽、芦苇、水莎草5个物种在3个时段皆有出现;而C区稗和芦苇的2个物种在3个时段皆有出现,D区岸带牛筋草和蔊菜等2个物种在3个时段皆有出现。4种生境土壤种子库的物种种类都由草本植物构成,且1年生草本所占比例与多年生草本所占比例相差不大。
|
|
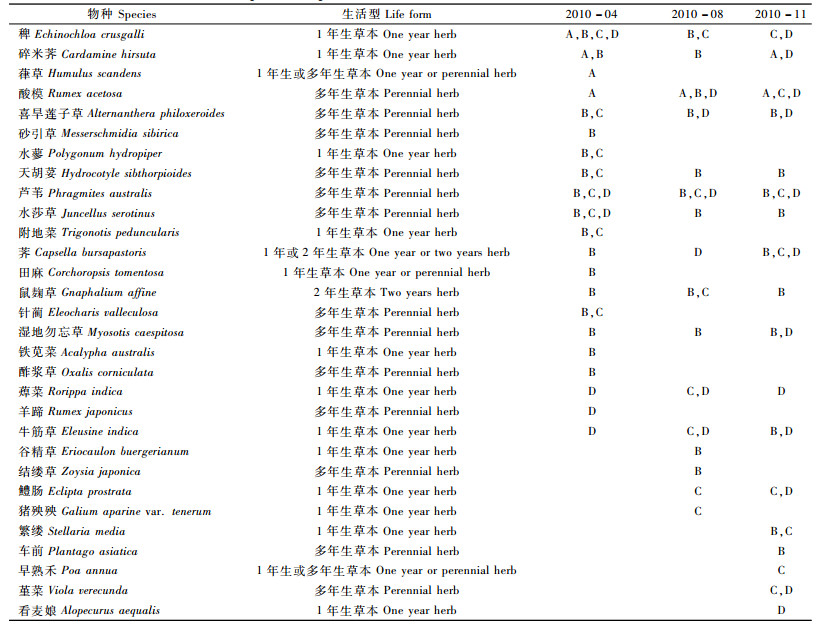
土壤种子库的平均物种数变动于3~5种之间。除了类型D在2010年11月达到最高(图 1)外,其他各植被类型的土壤种子库平均物种数均在4月达到最高。此外,除了11月4种不同覆被类型的土壤种子库平均物种数差异显著外(P < 0.05),其他不同采样时间和不同覆被条件下的土壤种子库平均物种数无显著差异。

|
图 1 不同采样时间各覆被类型土壤种子库的物种数 Fig.1 The number of species of soil seed banks in four types at different sampling time |
2010年4,8和11月种子库各月间的相似性如表 2,表明太湖岸带湿地土壤种子库物种组成有较大的季节差异。不同类型间群落空间相似性说明太湖岸带湿地不同植被类型对土壤种子库物种组成具有影响。另外,3个调查时段内的群落相似性有所差异,这表明种子库空间相似性程度也受季节的影响。
|
|

3个调查时段内土壤种子库密度的垂直分布由图 2可以看出,随着土层的加深,A,B,C,D 4种类型在不同时段的种子库密度垂直分布上大多呈下降趋势。其中,2010年4月调查时段内A,B,C,D 4种类型表层土壤种子库的储量分别占垂直分布的64.10%,60.44%,56.86%和69.05%。A,B,C,D区表层分别与中、下层显著差异(P<0.01)。其中,A区表层物种数占总物种数的75.0%;B区表层物种数占总物种数的87.5%;C区表层物种数占总物种数的87.5%;D区表层物种数占总物种数的83.3%。表层土壤不仅在种子库储量上占据多数,在物种数上也占据多数。在2010年8月和11月,类型B的表层土壤种子库数量较第2,3层低。这2个时段A,C,D 3种类型表层土壤种子库的储量分别占垂直分布的100%,71.43%,82.09%以及100%,57.14%,74.63%。

|
图 2 土壤种子库垂直分布 Fig.2 Vertical distribution of soil seed bank |
从整体看,4种植被类型中,太湖岸带湿地种子库2010年4月土壤种子库储量最高(图 3),为(6 280±1 731.20)粒·m-2;8月与11月种子储量相对较低,分别为(3 520±1 605.92)粒·m-2和(3 300±1 505.00)粒·m2。方差分析表明:4,8月与11月之间无显著差异(P>0.05)。针对4种不同植被覆被类型,除了类型D外,其余3种类型土壤种子库储量皆以4月最高,且随时间的变化明显;A,B,C 3种覆被类型种子库储量分别为(3 120±657.27)粒·m-2,(14 560±2 920.27)粒·m-2和(4 080±1 145.43)粒·m-2(图 4)。

|
图 3 土壤种子库储量的时间动态 Fig.3 The temporal dynamics of soil seed bank reserves |

|
图 4 不同覆被类型的太湖岸带湿地土壤种子库季节动态 Fig.4 Seasonal dynamics of lakeshore wetland seed bank in different cover types, Lake Tai |
研究检测出的太湖岸带湿地土壤种子库中的活力种子密度变动于(3 300~6 280)粒·m-2。这一密度值与湖北省和安徽省交界处的淡水湖泊——龙感湖湖滨沼泽的土壤种子库种子密度4 689粒·m-2较为接近(Yuan et al., 2007),但是这一密度值却远低于洞庭湖青山垸的(18 618~36 943)粒·m-2(侯志勇等, 2009)。对比不同研究结果的物种结构后发现,草本植物种子对差异贡献最大。从不同生活型物种组成来看,太湖岸带湿地1年生和多年生草本占所有萌发物种的90%以上。通常,不同生活型草本植物土壤种子库密度不同,在多年生草本植物占优势的区域,土壤种子库密度较小,而在1年生草本植物占优势的区域,土壤种子库密度较大,这主要是由于1年生草本植物种子数量较多,质量较轻;此外,多年生草本植物的种子产量比1年生草本植物低也是原因之一(Harper, 1977)。
太湖岸带湿地种子库的物种数量为30个,物种数量多于龙感湖湖滨的22个(Yuan et al., 2007),而少于鄱阳湖滨的35个(侯志勇等, 2009)。在本研究中,4种类型中土壤种子库共同物种只有稗,其生态位较宽,对干湿变化的环境具有较好的适应性。研究表明太湖岸带湿地土壤种子库在维持太湖岸带湿地地上植被植物多样性方面发挥着极其重要的作用,太湖大堤加固、加宽严重影响岸带湿地的状况,本研究为太湖岸带湿地的保护管理提供了技术支持。
目前,种子库的鉴定方法主要包括物理法和萌发法。其中,物理法主要包括漂浮浓缩法和网筛分选法(于顺利等, 2007)。物理法和萌发法都有各自的缺点,仍未有发现更好的方法。目前,萌发法广泛应用于湿地土壤种子库研究,其主要缺陷是萌发种子只是种子库中具有活力的一部分,部分不具活力的种子未被检出,从而低估了种子库的储量,将来在湿地土壤种子库研究中需要注意鉴定方法的选取。
太湖岸带湿地种子库储量的垂直分布大都表现为随土层的加深而减少。形成土壤种子库分布格局的机制可能与种子生产、扩散能力以及生境变化有关(徐洋等, 2009; 萧蒇等, 2011)。一般来说,土壤种子库具有较为明显的垂直分布格局,物种数量及种子库密度在表层土壤较高(Baldwin et al., 2001; Amiaud et al., 2004)。本研究中,3个不同时段内的种子库表层大都高于下层,同样符合上述规律。一般来说,种子掉落到土壤表层,经过动物践踏、搬运等多种方式形成二次分布而进入深层土壤中,多数种子储藏于土壤表层,有机会到达深层土壤的种子必然是少数(徐洋等, 2009; Baldwin et al., 2001)。原因是表层土壤温湿度较为适宜,通气性也较好,而随着土层的加深,土质变硬,透气性变差,且在种子深入土层的过程中,遇到的阻力也大(Capon et al., 2006; 葛斌杰等, 2010)。在2010年8月和11月,乔灌岸带的表层土壤种子库数量较第2,3层低。这可能与该区表层土壤中物种萌发特征有关,其萌发时间较早,在5,6月即完成萌发。
种子库的季节变化是由种子生产、扩散、续存和周转决定的(Lamont et al., 1991; Leck, 2003)。很多研究发现湿地土壤种子库的密度和物种组成具有明显的季节变化(王相磊等, 2003)。种子的输入和输出是决定土壤种子库季节变化的物质基础,而这与物种成熟节律和种子散落规律密切相关(于顺利等, 2007; 唐樱殷等, 2011)。本研究中除了天然芦苇湿地外,其余3种类型土层中种子库储量皆以4月最高,主要是由于4月承接了大量的上年秋熟散落的种子而使土壤种子库的物种数量和密度达到高值。天然芦苇湿地在2010年4月种子库储量较低可能与当地每年冬季对芦苇湿地进行火烧有关,当地政府每年冬季组织人力进行芦苇人工收割及火烧。火烧之后,由于温度过高使表层土壤中的种子失去活性。太湖岸带湿地种子库种子密度其次为夏季(8月),秋季(11月)最少。至8月,由于大部分种子在春季已经萌发,使土壤种子库的储量减少,种子库密度降低。至11月的秋季,虽然部分成熟种子逐步进入湿地土壤中,但是由于未经过“春化”过程,所以种子萌发率不高。同时,2010年9,10月降雨量偏多,有些种子成熟后因地表径流而流失,种子库密度表现为最低。此外,部分物种的种子成熟后推迟散落(Lamont et al., 1991),也使秋季土壤种子库种子密度受到影响。
| [] | 陈小华, 李小平, 张利权. 2007. 河道生态护坡技术的水土保持效益研究. 水土保持学报, 21(2): 32–35. |
| [] | 陈中义, 雷泽湘, 周进, 等. 2001. 梁子湖优势沉水植物冬季种子库的初步研究. 水生生物学报, 25(2): 52–158. |
| [] | 葛斌杰, 杨永川, 李宏庆. 2010. 天童山森林土壤种子库的时空格局. 生物多样性, 18(5): 489–496. |
| [] | 侯志勇, 谢永宏, 于晓英, 等. 2009. 洞庭湖青山垸退耕地不同水位土壤种子库特征. 应用生态学报, 20(6): 1323–1328. |
| [] | 李吉玫, 徐海量, 张占江, 等. 2008. 塔里木河下游不同退化区地表植被和土壤种子库特征. 生态学报, 28(8): 3626–3636. |
| [] | 李守淳, 刘文治, 刘晖, 等. 2011. 蚌湖湖滨带的土壤种子库特征. 植物科学学报, 29(2): 164–170. |
| [] | 李伟成, 盛海燕, 钟哲科, 等. 2007. 杭州西溪湿地中不同土地利用类型的种子库特性. 林业科学, 43(11): 163–169. DOI:10.3321/j.issn:1001-7488.2007.11.027 |
| [] | 刘贵华, 萧蒇, 陈漱飞, 等. 2007. 土壤种子库在长江中下游湿地恢复与生物多样性保护中的作用. 自然科学进展, 17(6): 741–747. |
| [] | 唐樱殷, 谢永贵, 余刚国, 等. 2011. 黔西北喀斯特土壤种子库季节动态及种子库对策. 生态学杂志, 30(7): 1454–1460. |
| [] | 王超, 王沛芳, 唐劲松, 等. 2003. 河道沿岸芦苇带对氨氮的削减特性研究. 水科学进展, 14(3): 311–317. |
| [] | 王相磊, 周进, 李伟, 等. 2003. 洪水湿地退耕初期种子库的季节动态. 植物生态学报, 27(3): 352–359. DOI:10.17521/cjpe.2003.0052 |
| [] | 王正文, 祝廷成. 2002. 松嫩草地水淹干扰后的土壤种子库特征及其与植被关系. 生态学报, 22(9): 1392–1398. |
| [] | 夏继红, 严忠民. 2006. 生态河岸带的概念及功能. 水利水电技术, 37(5): 14–17. |
| [] | 萧蒇, 刘文治, 刘贵华. 2011. 丹江口库区滩涂与入库支流植被与土壤种子库:水传播潜力探讨. 植物生态学报, 35(3): 247–255. |
| [] | 邢福, 王莹, 许坤, 等. 2008. 三江平原沼泽湿地群落演替系列的土壤种子库特征. 湿地科学, 6(3): 351–358. |
| [] | 徐洋, 刘文治, 刘贵华. 2009. 生态位限制和物种库限制对湖滨湿地植物群落分布格局的影响. 植物生态学报, 33(3): 546–554. |
| [] | 徐海量, 李吉玫, 王增如, 等. 2007. 塔里木河下游土壤种子库的空间分布特征分析. 水土保持学报, 21(6): 183–186. |
| [] | 叶春, 刘杰, 于海婵, 等. 2008. 东太湖3种沉水植物群落区底泥种子库与幼苗库. 生态环境, 17(3): 1091–1095. |
| [] | 于顺利, 陈宏伟, 郎南军. 2007. 土壤种子库的分类系统和种子在土壤中的持久性研究进展. 生态学报, 27(5): 2099–2108. |
| [] | Amiaud B, Touzard B. 2004. The relationships between soil seed bank, aboveground vegetation and disturbances in old embanked marshlands of Western France. Flora, 199(1): 25–35. DOI:10.1078/0367-2530-00129 |
| [] | Baldwin A H, Egnotovich M S, Clarke E. 2001. Hydrologic change and vegetation of tidal freshwater marshes: field, greenhouse, and seed bank experiments. Wetlands, 21(4): 519–531. DOI:10.1672/0277-5212(2001)021[0519:HCAVOT]2.0.CO;2 |
| [] | Capon S J, Brock M A. 2006. Flooding, soil seed bank dynamics and vegetation resilience of a hydrological variable desert floodplain. Freshwater Biology, 51(2): 206–223. DOI:10.1111/fwb.2006.51.issue-2 |
| [] | Harper J L. 1977. Population Biology of Plants. London, New York, San Francisco, Academic Press. |
| [] | Hu W, Sven E J, Zhang F B, et al. 2011. A model on the carbon cycling in Lake Taihu, China. Ecological Modelling, 222(16): 2973–2991. DOI:10.1016/j.ecolmodel.2011.04.018 |
| [] | Lamont B B, Maitre D C, Cowling R M, et al. 1991. Canopy seed storage in woody plants. The Botanical Review, 57(4): 277–317. DOI:10.1007/BF02858770 |
| [] | Leck M A. 2003. Seed bank and vegetation development in a created tidal freshwater wetland on the Delaware River, Trenton, New Jersey, USA. Wetlands, 23(2): 310–343. DOI:10.1672/9-20 |
| [] | Lu J, Wang H B, Pan M, et al. 2012. Using sediment seed banks and historical vegetation change data to develop restoration criteria for a eutrophic lake in China. Ecological Engineering, 39: 95–103. DOI:10.1016/j.ecoleng.2011.11.006 |
| [] | Thompson K, Grime J P. 1979. Seasonal variation in the seed bank of herbaceous species in ten contrasting habitats. Journal of Ecology, 67(3): 893–921. DOI:10.2307/2259220 |
| [] | Wang C, Wang P F. 2007. Hydraulic resistance characteristics of riparian reed zone in river. Journal of Hydrologic Engineering, 12(3): 267–272. DOI:10.1061/(ASCE)1084-0699(2007)12:3(267) |
| [] | Yuan L Y, Liu G H, Li W, et al. 2007. Seed bank variation along a water depth gradient in a subtropical lakeshore marsh, Longgan Lake, China. Plant Ecology, 189(1): 127–137. DOI:10.1007/s11258-006-9170-z |