 2012, Vol. 48
2012, Vol. 48文章信息
- 孙立方, 郭起荣, 王青, 冯云, 牟少华
- Sun Lifang, Guo Qirong, Wang Qing, Feng Yun, Mu Shaohua
- 毛竹花器官的形态与结构
- Flower Organs Morphology and Structure of Phyllostachys edulis
- 林业科学, 2012, 48(11): 124-129.
- Scientia Silvae Sinicae, 2012, 48(11): 124-129.
-
文章历史
- 收稿日期:2012-01-09
- 修回日期:2012-03-08
-
作者相关文章



2. 北京林业大学 北京 100083
2. Beijing Forestry University Beijing 100083
毛竹(Phyllostachys edulis)是我国特有的、最重要的经济竹种,也是我国分布最广、栽培历史长、栽培面积最大的竹种。据第7次全国森林资源清查报告显示,毛竹面积已达386.83万hm2,产生了巨大的经济、生态、社会效益。竹子作为禾本科(Gramineae)竹亚科(Bambusoideae)植物,长期以来多数以无性繁殖为主,不经常开花,即使像毛竹这样常见的种类,其生殖生物学领域研究少。近年见郭起荣等(2010)对其种子的研究,现报道对毛竹花的解剖学研究结果。
资料显示:1954年,浙江西天目山毛竹成片开花;1979,1983年,日本、美国毛竹局部大面积开花;1962—1980年,广西昭平、蒙山等毛竹数千公顷先后开花结实;1973年,江西兴国均福山林场毛竹大量开花;1975—1996年,广西荔浦、灵川、阳朔、兴安等地毛竹林不断开花(张文燕等,1989;Watanabe,1982;章日英等,1986;周芳纯,1998);进入2000年,广西毛竹不间断地零星开花。限于试验材料等因素的制约,仅见一些志书的描述,研究性文章少,描述不完整。
1 材料与方法试验材料2010,2011年6月采于广西桂北深山盛花期中的毛竹林中,海拔近千米,为中亚热带地区,年平均气温18~19 ℃,1月平均气温7~8 ℃,7月平均气温28~29 ℃,年平均降水量1 750~2 000 mm。
在全面调查开花林分基础上,选取生长良好的2,3年生植株中部花枝,随机剪取花序,进行结构解剖、测量、拍照。
于上午10点前,采集盛开小花和花药,分别放入FAA固定液,固定24 h以上,用作石蜡切片观察(李正理,1996),OLYMPUS CX31显微镜下观察、拍照。
将采集的新鲜花药置于干燥的硫酸纸上,阴干30~60 min,抖出花粉,放入干燥的离心管,密封,放入有冰袋的冰盒带回实验室。用镊子或毛笔取适量花粉放在洁净的载玻片上,滴加1,2滴蒸馏水,盖上盖玻片。将制备好的带有花粉的载玻片在显微镜(LEICA DM LB2)下,20倍物镜,随机选取5个视野,观察拍照,测量花粉的直径大小。
将经过FAA固定液处理的花药自然晾干,剥出花粉,直接涂散在导电双面胶带上,喷金镀膜,移入场发射环境扫描电子显微镜(美国FEI公司生产,XL30 ESEM FEG)下进行观察。试验数据均用SPSS16.0统计软件进行分析。
2 结果与分析 2.1 花序的形态结构花序是指许多花按一定的次序排列在茎轴上的方式,根据《中国植物志(第九卷第一分册)》和《中国树木志》,禾本科花序分为两大类型:一为单次发生花序(真花序);二为续次发生花序(假花序)。小穗是禾本科植物中组成花序的基本单位,由苞片、小花和小穗轴组成。竹类植物通常由少数花朵合成小穗,再由小穗聚合成圆柱状、总状、穗状或假圆锥等形状花序(江泽慧,2002)。毛竹为假花序类型,无限,以小穗组成整体的复穗状花序,基部托有2~7片逐渐增大的鳞片状苞片,不含小穗,多脱落;花序上部的佛焰苞4~13片,内各含有1个小穗(图 1)。随机抽样、统计20个毛竹花序,量得花序平均长(8.01±0.98)cm。

|
图 1 毛竹开花状及花序(花枝和花序类型) Fig.1 Flowering and inflorescence of P. edulis(flowering branch and inflorescence type) 1:花序轴Rachis;2:苞片Bract;3:佛焰苞Spathe. |
竹类植物的小穗,根据其基部的苞片或颖之腋内的潜伏芽的有无可以分为真小穗和假小穗。真小穗或称小穗无潜伏芽,而假小穗具潜伏芽或具前出叶(江泽慧,2002)。
桂北毛竹(图 2,3)每个花序平均包含10个小穗,为假小穗,4~13个不等,偏向一侧排列(扫帚状)。小穗基部有一苞片(佛焰苞),均长(3.56±0.43)cm,顶端锥状缩小叶,均长(1.04±0.13)cm;苞片内具一前出叶,均长(1.45±0.18)cm。小穗平均长(3.76±0.40)cm,花序轴节间距平均(0.48±0.07)cm,每小穗含小花数1~6朵不等,小穗轴具白色茸毛,顶端小花可孕或退化不孕。大于乔世义等(1984)在江西兴国研究报道的小穗长2.5~2.7 cm,含小花2~3枚,其中顶花不育,退化成针状,小穗轴节间短,长约0.2 cm等描述。这可能与当地气候(如积温等)对花器的生长发育相关。

|
图 2 毛竹穗状花序 Fig.2 Spicate inflorescence of P. edulis 1:假小穗Fake spikelet;2:花序轴Rachis(本样含9个小穗Specimen in picture with 9 spikelets). |

|
图 3 毛竹小穗(假小穗) Fig.3 Spikelet of P. edulis (false spikelet) 1:佛焰苞Spathe;2:前出叶Prophyll;3:小花Floret. |
典型竹类植物的小花包含颖片,外、内稃片,浆片(鳞被),花药,花丝,子房,花柱,柱头等。
通过解剖、统计30朵毛竹小花(图 5,6),计小花平均长(2.6±0.33)cm。

|
图 5 毛竹小花 Fig.5 Floret of P. edulis 1:雄蕊Stamen;2:雌蕊Pistil;3:最末级小穗轴The last rachilla;4:颖片Glume (1);5:内稃Palea;6:外稃Lemma. |

|
图 6 毛竹雌、雄蕊 Fig.6 Pistil and stamen of P. edulis 1:花药(3个)Anther(3);2:花丝(3个)Filament(3);3:浆片(3枚)Lodicule(3);4:子房Ovary;5:花柱Style;6:柱头(3裂)Stigma(3 dehiscence). |
颖片1~3枚,苞片状,薄革质,平均长(2.49±0.12)cm,顶端具锥状缩小叶,长0.3~0.5 cm,上下部边缘生长毛茸。
稃片2~3枚,外稃均长(2.58±0.14)cm,生有粗糙硬毛,先端锐尖;内稃稍短于外稃,平均长(2.33±0.12)cm,生有茸毛,背部具2脊,先端有齿裂。
浆片多数3枚,个别2枚,卵状披针形,白色透明膜质。
雄蕊3枚,均长(4.34±0.61)cm。花药淡黄色,平均长(1.26±0.10)cm,花丝平均长(2.98±0.52)cm。花药成熟后垂悬于伸出花丝而露出花外,平均伸出长(1.59±0.42)cm。同时花粉囊逐渐裂开纵缝,成熟花粉粒从裂缝散出。
通常,大多数被子植物的花药具有4个小孢子囊(花粉囊),少数具2个,它们由药隔组织分开。当花药达到成熟时,每一侧的2个小孢子囊往往药隔消失而连通。每个小孢子囊包含药室和由数层细胞组成的药壁(胡适宜,2005)。本研究表明毛竹花药为四室,每一花药由4个花粉囊组成,即四室,左右对称分开,中间以药隔相连(图 10),基部着生于花丝顶端,属于基着药(图 9),纵裂散粉(图 9,11)。与以前的研究描述相异:我国禾本科开创人耿义礼(1959)描述在禾本科花器官“每雄蕊具一纤细花丝与二室之花药。后者以基部附着花丝,但因呈深箭镞形,故视之似一背部附着而可转动”。中国科学院中国植物志编辑委员会(1996)描述为“具纤细的花丝与二室总裂开(稀可顶端孔裂)的花药,后者常以中部背着花丝顶端”;易同培等(2008)描述竹类小花“花药二室纵裂”;郑万钧(2004)描述为“花药丁字着生”;广西柳州地区贝江口林业试验站(1972)描述毛竹“花药二室”。而McClure(1983)报道,竹类“每一花药起先有四室,分成两对。当花粉粒成熟时,每一对的二室联成一囊。在花药伸出来并垂悬之后,花粉粒便通过顶孔从每一囊中释放出来,或者在有些竹子中,每一对花粉室在邻接边缘吻合处裂开一条长缝,花粉粒从这长裂缝中释放出来”。

|
图 9 毛竹雄蕊的扫描电镜照片 Fig.9 A stamen under scanning electron microscopy in P. edulis 1:花粉囊Clinandrium;2:花丝Filament;3:成熟花粉粒Mature pollen. |

|
图 10 毛竹花药横切面 Fig.10 Transverse section of anther of P. edulis 1:药隔Connective;2:花粉囊Clinandrium;3:药室Locule;4:花粉Pollen. |

|
图 11 毛竹成熟花药横切面 Fig.11 Transverse section of mature anther of P. edulis 1:裂缝(纵裂)Fissure (in lengthway). |
雌蕊1枚,子房1枚,上位,一室(图 8);胚珠1枚,倒生;柱头羽状三裂,属于长花柱型。花柱平均长(1.19±0.08)cm,柱头平均长(1.70±0.17)cm。开花时稃片张开,柱头伸出稃外,平均露出长(0.95±0.23)cm。没有发现Zhang等(1986)报道的美国毛竹因柱头外露短,影响授粉的问题。

|
图 8 毛竹子房纵切面 Fig.8 Longitudinal section of a ovary of P. edulis 1:胚珠Ovule;2:珠心Nucellus;3:珠被Integument;4:合点Chalaza;5:二核胚囊Two-nucleus embryo sac;6:珠柄Funicle. |
当花药成熟散粉后逐渐皱缩干枯,随后脱落。随机抽取3个小花,测定9个花药,花药的平均花粉数为(15 807±1 514)个。解剖中还发现,除了稃片和浆片外,外稃内、内稃背基部有一细小硬质丝尖状物,短直、稍软如芒,皇针状(图 4-2,5-3,7-6),长度大于1 cm,随着毛竹谷粒的发育成熟,最终隔内稃压在线状种脐腹沟,几与果实同长。

|
图 4 毛竹花小穗轴 Fig.4 Rachilla of P. edulis 1:小穗轴Rachilla;2:最末级小穗轴The last rachilla;3:顶端小花Top floret. |

|
图 7 毛竹小花横切面 Fig.7 Transverse section of a floret of P. edulis 1:外稃Lemma;2:内稃Pelea;3:浆片Lodicule;4:花药Anther;5:子房Ovary;6:最末级小穗轴The last rachilla. |
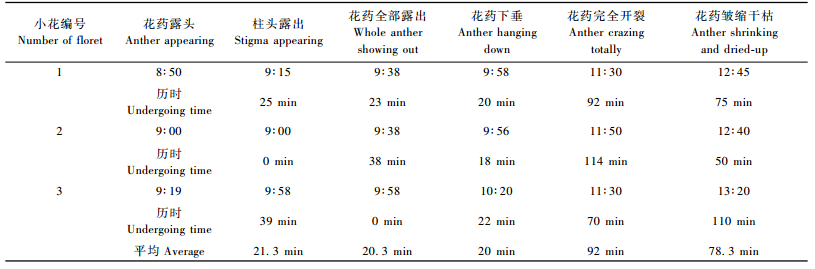
毛竹小花多以上午8:30—10:30,下午3:00—4:30开放,以上午露水多,湿度大,利于稃片吸胀、外张而开放为主。此2段时间试验地内可听、可见大量蜜蜂来访。
选择连续晴天上午观察30朵小花得知,从稃片张开,花药露头到花药皱缩干枯持续时间多为1~2 h,亦有长达5 h的,但均不及乔士义(1984)报道的开放持续时间约10 h。
|
|
对毛竹花粉的形态、结构尚未见报道。本研究在显微镜下测得36粒花粉(图 12),其平均直径为61.00 μm,最大直径为68.25 μm,最小51.20 μm,大于林树燕(2009)报道的异叶苦竹(Arundinaria simonii f. albostriatus)27.86 μm、鹅毛竹(Shibataea chinensis)40.22 μm的花粉直径。也大于刘志玲等(1999)报道的水稻(Oryza sativa)、玉米(Zea mays)、狼尾草(Pennisetum alopecuroides)花粉的直径。环境扫描电镜下观察花粉为近球形(图 13),外壁纹饰为细颗粒状,具一个圆形的萌发孔(图 14),孔周围有一圈孔环,外孔径为8.69 μm,内孔径为4.52 μm。毛竹花粉形态与鹅毛竹、异叶苦竹相近,而显著区别于水稻、玉米、狼尾草。

|
图 12 显微镜下的花粉 Fig.12 Pollens under microscope |

|
图 13 环境扫描电镜下的花粉 Fig.13 Pollens under the environmental scanning electron microscope |

|
图 14 花粉萌发孔 Fig.14 Germinal aperture of pollen 1:萌发孔Germinal aperture;2:孔环Circle. |
作为假花序类型的毛竹由穗状花序之小穗组成复穗状花序,平均长8.01 cm;每个花序平均含10个假小穗,小穗平均长3.76 cm,有颖片1~3枚,花序轴节间距均长0.48 cm。每小穗含小花数1~6朵不等,小花平均长2.6 cm。小花含稃片2~3枚,浆片多数3枚,个别2枚。雄蕊3枚,均长4.34 cm,花丝均长2.98 cm,花药均长1.26 cm,为四室,基着药,完全开放时平均伸出稃片外1.59 cm,一花药平均花粉数量15 807个,花药露头至花粉散布历时1~2 h。雌蕊1枚,子房1枚,上位,一室;胚珠1枚,倒生;花柱均长1.19 cm,柱头平均长1.70 cm,开放完整时平均露稃片外长0.95 cm。这可能与本试验地为多年的毛竹开花主要区域——桂北的积温等气候因素引起的生长发育相关。本研究所得数据填补了毛竹花器官形态解剖科学数据。
通过环境扫描电镜观察毛竹花粉平均直径为61.00 μm,近球形,外壁纹饰为细颗粒状,具一个圆形的萌发孔,萌发孔外孔径为8.69 μm,内孔径为4.52 μm,孔周围有一圈孔环等特征。
近年,其他竹类的花器解剖报道中,较少有对花药分室、着生和开裂散粉方式的描述,仅见黄新红(2008)报道巨龙竹(Dendrocalamus sinicus)花药四室,其他如林树燕(2009)发现鹅毛竹为基着药,刘静(2008)则观察到月月竹(Chimonobambusa sichuanensis)为丁字药且孔裂散粉,张文燕等(1992)也指出五月季竹(Phyllostachys bambusoides)花药是孔裂散粉。而林树燕(2009)、何其江等(2003)、邢新婷等(2005)、董文渊等(2001)、唐赛春等(2006)、李晓芬等(2009)、彭晟等(2006)、王小红等(2009)对水竹(Phyllostachys heteroclada)、雷竹(Phyllostachys praecox cv.prevernalis)、紫竹(Phyllostachys nigra)、麻竹(Dendrocalamus latiflorus)、黄竹(Dendrocalamus ronganensis)、慈竹(Neosinocalamus affinis)、筇竹(Qiongzhuea tumidinoda)、异叶苦竹、云龙箭竹(Fargesia papyrifera)、元江箭竹(Fargesia yuanjiangensis)等的花器解剖均没有对其花药分室、着生方式和开裂散粉方式进行描述。从本研究结果看,毛竹花药为四室,左右对称分开,基着药(图 9),纵裂散粉,而过去报道的“二室”可能指因成熟花药药隔消失而引起的药室合并,也可能因未借助显微设备,导致观察不确切。
| [] | 蔡春菊, 刘凤, 郭起荣, 等. 2010. 毛竹种子种质保存对含水量的响应. 江西农业大学学报, 32(2): 312–317. |
| [] | 董文渊, 黄宝龙, 谢泽轩, 等. 2001. 筇竹开花结实特性的研究. 南京林业大学学报:自然科学版, 25(6): 30–32. |
| [] | 耿以礼. 1956. 中国主要植物图说·禾本科. 北京, 科学出版社. |
| [] | 广西柳州地区贝江口林业试验站. 1972. 毛竹种子育苗和造林. 南宁, 广西人民出版社. |
| [] | 郭起荣, 任立宁, 牟少华, 等. 2010. 毛竹种质分子鉴别ARAP、AFLP、ISSR联合分析. 江西农业大学学报, 32(5): 982–986, 991. |
| [] | 何奇江, 汪奎宏, 华锡奇, 等. 2003. 雷竹花穗和花器的观察研究. 浙江林业科技, 23(2): 10–11, 15. |
| [] | 胡适宜. 2005. 被子植物生殖生物学. 北京, 高等教育出版社. |
| [] | 黄大勇. 2009. 广西竹类植物. 南宁, 广西科学技术出版社. |
| [] | 黄新红. 2008. 巨龙竹生殖生物学研究. 西南林学院硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10677-2009095570.htm |
| [] | 江泽慧. 2002. 世界竹藤. 沈阳, 辽宁科技出版社. |
| [] | 李蓉, 郭起荣, 曾炳山, 等. 2008. 毛竹种子"以芽繁芽"组培快繁初步研究. 世界竹藤通讯, 6(6): 9–13. |
| [] | 李晓芬, 林新春, 时燕, 等. 2009. 紫竹开花生物学特性观察及花粉生活力测定. 林业科技开发, 23(6): 64–67. |
| [] | 李正理. 1996. 植物组织制片. 北京, 北京大学出版社. |
| [] | 林树燕. 2009. 鹅毛竹和异叶苦竹的生殖生物学研究. 南京林业大学博士学位论文. http://cdmd.cnki.com.cn/article/cdmd-10298-2009180982.htm |
| [] | 刘静. 2008. 月月竹开花生物学研究. 南京林业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10298-2008112979.htm |
| [] | 刘志玲, 付建华, 陈良碧, 等. 1999. 三种禾本科植物花粉的结构比较. 湖南师范大学自然科学学报, 22(1): 70–74. |
| [] | 彭晟, 杨汉奇, 李德铢. 2006. 竹亚科箭竹属两种植物花序的补充描述. 云南植物研究, 28(3): 257–258. |
| [] | 乔士义. 1984. 毛竹开花生物学特性观察. 竹类研究(2): 20–24. |
| [] | 唐赛春, 夏念和, 林汝顺. 2006. 融安黄竹小穗和小花的形态发育. 植物分类学报, 44(5): 551–558. |
| [] | 王小红, 郭起荣, 周祖基. 2009. 水竹和慈竹开花代谢关键因子主成分分析. 林业科学, 45(10): 158–162. DOI:10.11707/j.1001-7488.20091028 |
| [] | 温太辉. 1993. 中国竹类彩色图鉴. 台北, 淑馨出版社. |
| [] | 邢新婷, 傅懋毅, 肖贤坦. 2005. 麻竹开花生物学特性观察及控制授粉的初步研究. 北京林业大学学报, 27(6): 103–107. |
| [] | 易同培, 史军义, 马丽莎. 2008. 中国竹类图志. 北京, 科学出版社. |
| [] | 张文燕, 马乃训. 1989. 竹类植物花期生物学特性. 林业科学研究, 2(6): 596–600. |
| [] | 郑万钧. 2004. 中国树木志. 北京, 中国林业出版社.. |
| [] | 周芳纯. 1998. 竹类植物开花结实. 竹类研究(1): 93–101. |
| [] | 中国科学院中国植物志编辑委员会. 1996. 中国植物志. 第9卷第1分册. 北京, 科学出版社. |
| [] | McClure F A. 1983. 洪效训, 译. 竹子的生殖相(下). 竹子研究汇刊, 2(2): 119-136. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=zzyj198302009&dbname=CJFD&dbcode=CJFQ |
| [] | Watanabe M. 1982. Flowering, seeding, germination and flowering periodicity of Phyllostachys pubescens. Jap For Soc, 64(3): 107–111. |
| [] | Zhang R Y(章日英), Cooper S G, Hansken J, et al.1986.Research on the flowering and propagation of Phyllostachys pebescens in America.Journal of Bamboo Research(竹子研究汇刊), 5(2): 44-52. http://en.cnki.com.cn/article_en/cjfdtotal-zzyj198602004.htm |