 2012, Vol. 48
2012, Vol. 48文章信息
- 崔鸿侠, 肖文发, 潘磊, 黄志霖, 王晓荣, 庞宏东
- Cui Hongxia, Xiao Wenfa, Pan Lei, Huang Zhilin, Wang Xiaorong, Pang Hongdong
- 神农架巴山冷杉林土壤碳储量特征
- Characteristics of Soil Carbon Storage of Abies fargesii Forest in Shennongjia
- 林业科学, 2012, 48(11): 107-111.
- Scientia Silvae Sinicae, 2012, 48(11): 107-111.
-
文章历史
- 收稿日期:2012-01-04
- 修回日期:2012-10-14
-
作者相关文章



2. 湖北省林业科学研究院 武汉 430075
2. Hubei Academy of Forestry Wuhan 430075
国内外学者对土壤有机碳储量及其影响因素进行了大量研究,取得了一定成果(Eswaran et al., 1993;Scott et al., 2002;王绍强等,2000;解宪丽等,2004),但由于土壤有机碳库组成的复杂性和影响因素的多样性,目前国内外对土壤有机碳库的储量、分布等研究仍显不足(吴建国等,2002;Torn et al., 1997;Jackson et al., 2000)。
神农架拥有在当今世界中纬度地区保持完好的亚热带森林生态系统,但在这一区域内,至今没有开展森林生态系统植被及土壤碳储量的研究,因此研究神农架地区典型森林类型土壤碳储量,有助于丰富对我国不同区域碳储量特征的认识。
巴山冷杉(Abies fargesii)是我国亚热带地区暗针叶林的主要树种之一,也是湖北神农架亚高山地区最重要的建群种。20世纪60年代,由于人为的大量砍伐,神农架自然保护区中大片的巴山冷杉原始林消失,取而代之的是一丛丛低矮的灌木和草甸。目前对神农架巴山冷杉林的研究主要集中在种群结构、分布格局和植被演替等方面(蒋雪琴等,2009;于倩等,2008;李景侠等,2001;李文漪,1991)。本研究在神农架自然保护区内选择巴山冷杉原始林、20世纪60年代末原始林砍伐后自然恢复形成的巴山冷杉次生林以及砍伐后第2年栽植的巴山冷杉人工林3种森林类型,研究不同起源巴山冷杉林的土壤有机碳储量及其影响因素,为我国土壤碳库的准确估算做出贡献,并为提高神农架地区土壤碳库管理水平提供理论依据。
1 研究区概况研究区位于神农架自然保护区(110°03′05″—110°33′50″E,31°21′20″—31°36′20″N),保护区总面积70 467 hm2。区内山势高大、山坡陡峻、河谷深切,山峰海拔多在1 500 m以上。保护区内地貌类型复杂,除主要的山地地貌外,还有流水地貌、喀斯特地貌和第四纪形成的冰川地貌。保护区气候属于北亚热带向暖温带过渡气候,年平均气温11.0~12.2 ℃,极端最低气温-21.2 ℃,极端最高气温40.5 ℃。多年平均日照率42%,空气相对湿度为73%~75%。平均年降水量800~2 500 mm,主要集中在夏季,从总体上看属于湿润气候带。
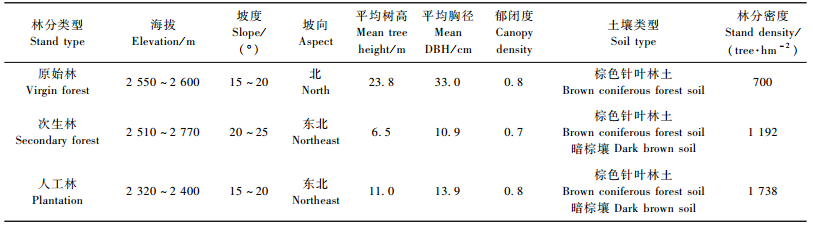
保护区内巴山冷杉原始林主要分布在海拔2 500 m以上,原始林内多数林木年龄超过100年,少数林木达到1 000年。乔木层唯一优势树种是巴山冷杉,林下灌木层物种较少,以箭竹(Fargesia spathacea)为主,草本层主要有赤车(Pellionia radicans)和山酢浆草(Oxalis griffithii)等。巴山冷杉次生林主要是原始林皆伐后未经人为干扰而天然更新形成,多分布在交通条件差,造林不方便地区。乔木层以巴山冷杉为主,混交有少量红桦(Betula albo-sinensis)和紫枝柳(Salix heterochroma),下木层物种丰富,主要有箭竹、五尖槭(Acer maximowiczii)、无隐悬钩子(Rubus inopterus)、三脉紫菀(Aster ageratoides)和草地早熟禾(Poa pratensis)等。巴山冷杉人工林为原始林皆伐后造林形成,林龄达40年。采伐后主要进行了剩余物清理等工作,造林整地方式为挖穴,初植密度为2 500株·hm-2,造林后采取封育措施。人工林乔木层以巴山冷杉为主,混交少量红桦和山杨(Populus davidiana),下木层主要有五尖槭、紫花卫矛(Euonymus porphyreus)和尾萼蔷薇(Rosa caudata)等。
2 研究方法 2.1 样地设置与土样采集2011年9月选择巴山冷杉原始林、次生林和人工林3种类型,各设置3块20 m×20 m样地,共计9块样地。在各样地内均匀布设5块2 m×2 m的灌木样方,灌木样方内再取1 m×1 m草本植物样方,进行乔木层每木调查和灌草层群落调查。并分别在每个样地按S形布设5个采样点,按照0~10,10~20,20~40和40~60 cm土层用直径为5 cm的土钻取出完整的土芯,将5个采样点同一层的土样混合均匀,留取1 kg土样自然风干,用于测定土壤有机碳含量。同时用环刀法测定土壤密度。样地基本情况见表 1。
|
|
土壤有机碳含量测定采用重铬酸钾氧化外加热法(刘光崧,1996);土壤碳储量计算公式为CSO=C×D×E×(1-G)/10,CSO为土壤碳储量(t·hm-2),C为土壤有机碳含量(g·kg-1),D为土壤密度(g·cm-3),E为土层厚度(cm),G为直径>2 mm的石砾所占的体积比例(%)。
2.3 凋落物现存量测定2011年11月在9块样地内各设置3块0.5 m×0.5 m的小样方,按照未分解、半分解和已分解分别收集样方内的全部凋落物,带回实验室称取鲜质量。再从样方内收集的枯枝落叶中取一定量的样品,采用恒温烘干法,测定其含水量,最后把鲜质量换算为干物质生物量。
2.4 数据分析数据分析采用Microsoft Excel和SAS 9.2统计软件进行数据处理分析。
3 结果与分析 3.1 凋落物现存量如表 2所示,不同起源巴山冷杉林未分解层凋落物现存量变化范围为4.69~6.24 t·hm-2,半分解层为5.65~21.89 t·hm-2,已分解层为42.62~81.11 t·hm-2,凋落物总量变化范围为53.03~97.15 t·hm-2,算术平均值为77.80 t·hm-2,大小顺序为原始林>人工林>次生林。同一起源林分不同分解层比较,凋落物现存量大小均表现为已分解层>半分解层>未分解层。差异显著性分析结果表明,不同起源林分凋落物半分解层、已分解层差异均达到显著性水平(P<0.05)。
|
|

如表 3所示,土壤有机碳含量在不同起源的巴山冷杉林中均随着土层的加深而减小,差异显著性分析结果表明,人工林土壤有机碳含量在各层之间均有显著性差异。在不同土层中,原始林土壤有机碳含量均大于次生林和人工林,而次生林和人工林土壤有机碳含量在不同土层中大小关系并不固定。
|
|

利用公式计算的土壤有机碳储量结果见表 4。原始林、次生林和人工林0~60 cm土层总碳储量分别为271.93,218.29和219.80 t·hm-2。在不同土层中,原始林土壤有机碳储量数值上均大于次生林和人工林。由于土壤碳储量的计算受土壤有机碳含量、土壤密度和土层厚度等多个因子的影响,因此不同土层之间的土壤碳储量变化规律与土壤碳含量的变化存在差异性。
|
|

采用Shannon-Wiener物种多样性指数,计算得出神农架巴山冷杉原始林、次生林和人工林林下灌木层物种多样性指数分别为0.63,1.74和1.71,草本层物种多样性指数分别为0.70,1.59和1.52。将灌木层和草本层物种多样性指数分别与0~60 cm土层总碳储量进行回归分析,结果见图 1。从图 1可知,神农架巴山冷杉林地土壤总碳储量与灌木层和草本层的物种多样性均呈显著的负相关关系(P<0.05)。

|
图 1 土壤碳储量与物种多样性指数的关系 Fig.1 Relationship between soil carbon storage and Shannon-Wiener species diversity index |
对不同土层的土壤碳储量与凋落物现存量分别进行回归分析,结果如图 2所示。从图 2可知,凋落物现存量与0~10,10~20和20~40 cm土层土壤碳储量存在显著的正相关关系(P<0.05),而与40~60 cm土层土壤碳储量相关性不显著(P>0.05)。

|
图 2 土壤碳储量与凋落物现存量的关系 Fig.2 Relationship between soil carbon storage and litter storage |
本研究结果显示,在同一起源的林分中,土壤有机碳含量随着土层的加深而减小,表现出明显的表聚性,这与以前的研究结果一致(常宗强等,2008;田大伦等,2011)。在不同起源的不同层次土壤中,均表现出原始林的土壤有机碳含量和储量大于次生林和人工林,这种差异可能与原始林林下凋落物存储量大、向土壤输入的有机碳多、林分形成时间较次生林和人工林长、林分郁闭早、林地土壤温度低和人为干扰极少等原因有关。神农架巴山冷杉原始林、次生林和人工林0~60 cm土层总碳储量分别为271.93,218.29和219.80 t·hm-2,神农架巴山冷杉林土壤碳储量远大于全国土壤平均碳储量(96.0 t·hm-2)(于东升等,2005)。巴山冷杉原始林土壤碳储量大于全国棕色针叶林土平均碳储量(247.4 t·hm-2),巴山冷杉次生林和人工林土壤碳储量虽小于全国棕色针叶林土碳储量平均值,但均大于全国暗棕壤平均碳储量(187.6 t·hm-2)(于东升等,2005)。可见,神农架巴山冷杉林土壤在全国土壤碳库中作出了重大贡献。
与巴山冷杉原始林碳储量相比较,森林砍伐后形成的次生林和人工林碳储量分别下降19.73%和19.17%。森林砍伐一方面带走了大量植物残体,减少了土壤有机质的输入量,降低了土壤有机碳储存量;另一方面使土壤失去了植物保护,导致水土流失,使大量的土壤有机碳随之流失。此外,森林砍伐还改变土壤的温度、湿度,影响土壤中微生物的数量、种类及其活性,对土壤有机碳的矿化分解有很大影响。森林砍伐后的不同恢复措施对土壤有机碳的影响不同,森林转化为农田,通常土壤碳损失25%~40%,耕作层的损失量最大,一般可达40%;森林转为草地和轮作地,土壤碳分别损失20%和18%~27%(周广胜等,2002)。此外森林砍伐后的恢复年限也是影响土壤有机碳的重要因素,张鼎华等(2002)对亚热带常绿阔叶林和杉木林砍伐后土壤肥力变化进行了研究,发现皆伐106天后土壤有机碳分别降低34.7%和24.4%;Brown等(1984)研究表明砍伐森林或改变林地利用现状都会造成前20~30年内多至20%~50%的有机碳损失,而随着恢复年限的增长土壤有机碳会有一定程度的积累。本研究区皆伐林地在恢复40年后土壤有机碳储量降低近20%,这与前人的研究结果具有较好的一致性。可见,森林砍伐后,其土壤有机碳含量和碳储量将减少,但减少的程度取决于紧接着的恢复措施及年限。因此,减少人为活动的负面干扰,增强森林保护,有利于维持和增加土壤碳储量,对于缓解大气中CO2浓度具有重要意义。
植物凋落物分解是土壤有机质输入的主要来源,是土壤呼吸的一个重要组成部分(Bowden et al., 1993;Sulzman et al., 2005)。王光军等(2009a;2009b)研究了去除和添加凋落物对枫香(Liquidambar formosana)、樟树(Cinnamomum camphora)和马尾松(Pinus massoniana)等森林类型土壤呼吸的影响;郭胜利等(2009)研究了黄土区不同植被类型下凋落物对土壤有机碳和可溶性碳的影响,结果均表明凋落物是影响土壤碳的一个重要因素。本次研究结果发现凋落物现存量与0~10,10~20和20~40 cm土层土壤碳储量显著正相关(P<0.05),而与40~60 cm土层土壤碳储量相关性不显著,这与郭胜利等(2009)的研究结果一致,可见凋落物对表层土壤有机碳的影响大于下层土壤。
目前,国外许多学者对草地生态系统植被碳储量与物种多样性的关系开展了研究,并普遍认为植物物种多样性的减少意味着生态系统固定CO2的能力降低,植被碳储量随着物种多样性的降低而减少(Hector et al., 1999;Loreau,2000;Craine et al., 2003;Hooper et al., 2005)。而Enquist等(2001)发现碳储量与物种多样性之间没有显著的相关关系。本研究发现神农架巴山冷杉林灌木层和草本层的物种多样性与土壤碳储量均有显著的负相关关系。植物物种多样性对土壤碳储量的影响体现在2方面:一方面物种多样性高的植物群落对空间、光照和养分利用率高,从而使植物的生物量最大化,地上生物量高意味着土壤碳输入量大,有利于土壤碳积累;另一方面植物物种多样性通过影响土壤微生物群落的多样性,从而影响土壤有机质的分解。不同植被类型植物物种多样性与土壤碳储量的关系,还存在不确定性,值得进一步研究。
| [] | 常宗强, 冯起, 司建华, 等. 2008. 祁连山不同植被类型土壤碳贮量和碳通量. 生态学杂志, 27(5): 681–688. |
| [] | 郭胜利, 马玉红, 车升国, 等. 2009. 黄土区人工与天然植被对凋落物量和土壤有机碳变化的影响. 林业科学, 45(10): 14–18. DOI:10.11707/j.1001-7488.20091003 |
| [] | 蒋雪琴, 刘艳红, 赵本元. 2009. 湖北神农架地区巴山冷杉种群结构特征与空间分布格局. 生态学报, 29(5): 2211–2218. |
| [] | 李景侠, 张文辉. 2001. 巴山冷杉种群结构及空间分布格局的研究. 西北农林科技大学学报, 29(5): 115–118. |
| [] | 李文漪. 1991. 神农架巴山冷杉林花粉与植物关系及其森林植被之演替. 地理学报, 46(2): 186–194. |
| [] | 刘光崧. 1996. 中国生态系统研究网络观测与分析标准方法——土壤理化分析与剖面描述. 北京, 中国标准出版社. |
| [] | 田大伦, 王新凯, 方晰, 等. 2011. 喀斯特地区不同植被恢复模式幼林生态系统碳储量及其空间分布. 林业科学, 47(9): 7–14. DOI:10.11707/j.1001-7488.20110902 |
| [] | 王光军, 田大伦, 闫文德, 等. 2009a. 去除和添加凋落物对枫香和樟树林土壤呼吸的影响. 生态学报, 29(2): 643–652. |
| [] | 王光军, 田大伦, 闫文德, 等. 2009b. 马尾松林土壤呼吸对去除和添加凋落物处理的响应. 林业科学, 45(1): 27–30. |
| [] | 王绍强, 周成虎, 李克让., 等. 2000. 中国土壤有机碳库及空间分布特征分析. 地理学报, 55(5): 533–544. DOI:10.11821/xb200005003 |
| [] | 吴建国, 张小全, 王彦辉, 等. 2002. 土地利用变化对土壤物理组分中有机碳分配的影响. 林业科学, 38(4): 19–29. DOI:10.11707/j.1001-7488.20020404 |
| [] | 解宪丽, 孙波, 周慧珍, 等. 2004. 中国土壤有机碳密度和储量的估算与空间分布分析. 土壤学报, 41(1): 35–43. DOI:10.11766/trxb200301140106 |
| [] | 于东升, 史学正, 孙维侠, 等. 2005. 基于1:100万土壤数据库的中国土壤有机碳密度及储量研究. 应用生态学报, 16(12): 2279–2283. DOI:10.3321/j.issn:1001-9332.2005.12.011 |
| [] | 于倩, 谢宗强, 熊高明, 等. 2008. 神农架巴山冷杉林群落特征及其优势种群结构. 生态学报, 28(5): 1931–1941. |
| [] | 张鼎华, 范少辉. 2002. 亚热带常绿阔叶林和杉木林皆伐后林地土壤肥力的变化. 应用与环境生物学报, 8(2): 115–119. |
| [] | 周广胜, 王玉辉, 蒋延玲, 等. 2002. 陆地生态系统类型转变与碳循环. 植物生态学报, 26(2): 250–254. |
| [] | Bowden R D, Nadelhoffer K J, Boone R D, et al. 1993. Contributions of aboveground litter, belowground litter and root respiration to total soil respiration in a temperate hardwood forest. Canadian Journal of Forest Research, 23(7): 1402–1407. DOI:10.1139/x93-177 |
| [] | Brown S, Lugo A E. 1984. Biomass of tropical forests: a new estimate based on forest volumes. Science, 223(4642): 1290–1293. DOI:10.1126/science.223.4642.1290 |
| [] | Craine J M, Reich P B, Tilman D, et al. 2003. The role of plant species in biomass production and response to elevated CO2 and N. Ecology Letters, 6(7): 623–625. DOI:10.1046/j.1461-0248.2003.00467.x |
| [] | Enquist B J, Niklas K J. 2001. Invariant scaling relations across tree-dominated communities. Nature, 410(6829): 655–660. DOI:10.1038/35070500 |
| [] | Eswaran H, Vander B E, Reich P. 1993. Organic carbon in soils of the world. Soil Science Society of America Journal, 57(1): 192–194. DOI:10.2136/sssaj1993.03615995005700010034x |
| [] | Hector A, Schmid B, Beierkuhulein C, et al. 1999. Plant diversity and productivity experiments in European grasslands. Science, 286(5442): 1123–1127. DOI:10.1126/science.286.5442.1123 |
| [] | Hooper D U, Chapin I F, Ewel J J, et al. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs, 75(1): 3–35. DOI:10.1890/04-0922 |
| [] | Jackson R B, Schenk H J, Jobbagy E G, et al. 2000. Belowground consequences of vegetation change and their treatment in models. Ecological Applications, 10(2): 470–483. DOI:10.1890/1051-0761(2000)010[0470:BCOVCA]2.0.CO;2 |
| [] | Loreau M. 2000. Biodiversity and ecosystem functioning: recent theoretical advances. Oikos, 91(1): 3–17. DOI:10.1034/j.1600-0706.2000.910101.x |
| [] | Scott N A, Tate K R, Giltrap D J, et al. 2002. Monitoring land-use change effects on soil carbon in New Zealand: quantifying baseline soil carbon stocks. Environmental Pollution, 116(S1): 167–186. |
| [] | Sulzman E W, Brant J B, Bowden R D, et al. 2005. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry, 73(1): 231–256. DOI:10.1007/s10533-004-7314-6 |
| [] | Torn M S, Trunbore S E, Chadwick O A, et al. 1997. Mineral control of soil organic carbon storage and turnover. Nature, 389(6647): 170–173. DOI:10.1038/38260 |