 2012, Vol. 48
2012, Vol. 48文章信息
- 王静, 梁军, 焦一杰, 张星耀
- Wang Jing, Liang Jun, Jiao Yijie, Zhang Xingyao
- 杨树人工林林木个体大小比数与溃疡病发生程度的关系
- Relationships between Neighborhood Comparison of Short-Rotation Poplar Plantations and Canker Disease Incidence
- 林业科学, 2012, 48(11): 57-62.
- Scientia Silvae Sinicae, 2012, 48(11): 57-62.
-
文章历史
- 收稿日期:2012-03-05
- 修回日期:2012-04-24
-
作者相关文章



杨树(Populus)人工林在我国的种植规模已超过1 000万hm2。长期以来,我国杨树生产受到多种病虫害的严重制约,其中杨树溃疡病(Botryosphaeria dothidea)作为典型的寄主主导性病害,其病原广泛地存在于自然界及潜伏侵染于树木体内,严重影响树木生长并导致寄主死亡。该病主要分布在我国东北、华北、西北地区(张星耀等,2003;黄烈健等,2003)。气候和立地生态逆境是这类病害发生的主要诱因,当逆境生态因子导致树木生长势下降至某一个阈值时,病原-寄主复合体开始病理过程,影响树木生长,造成树木死亡。由于林木病害的发生与林分结构密切相关(Larsson,1989),而空间结构决定树木之间的竞争及其空间生态位,在很大程度上决定林分的稳定性、发展的可能性和经营空间的大小。对于单一树种组成的杨树人工林而言,优化林分空间结构,提高树木生长势,增加抵抗胁迫能力,是防治杨树溃疡病的有效途径。
林分的空间结构是指林木的分布格局及其属性在空间上的排列方式,反映了森林群落内物种的空间关系(Mason et al., 1995;Ferris et al., 1999;Pommerening,2002)。传统上采用的树种组成、胸径、树高等林分结构因子,虽然在一定程度上表达了林分的均值信息,但这些结构因子大多不含空间结构信息。近年来对林分空间结构的研究中提出了多种描述空间结构的参数,如混交度、角尺度和大小比数(惠刚盈等,1999;2001;2003;雷相东等,2002)。目前,用这3个参数描述空间结构在北方森林群落结构中研究较多(Hu et al., 2003;汤孟平等,2004;张群等,2003)。对于人工纯林的空间结构,描述树种或单株的生长优势状态的大小比数参数得以广泛应用(郝云庆等,2006)。其中树种的大小比数用来表示该树种在任意一可度量因子上,与其周围最近几株相邻木的空间大小配置。大小比数能够准确地量化参照树与相邻木的大小关系,从而使在空间结构参数基础上人工重复复杂的林分结构成为可能。因此,研究林木之间的空间关系以及这种空间结构对树木个体病害发生的影响,对指导以营林措施为导向的病害生态控制具有重要的指导意义。
本研究采用林木的胸径、冠幅和树高的大小比数作为参数,以7年生欧美杨107 (P.×euramericana cv. Neva)、欧美杨108 (P.×euramericana cv.Guariento)和欧美杨DN113 (P.×euramericana cv. DN113)为研究对象,对林木不同指标个体优势度与杨树溃疡病发病程度的关系进行分析;以欧美杨107为例,分析不同密度下不同指标大小比数与杨树溃疡病发病程度的关系,为杨树溃疡病预防提供可释性强的空间结构信息支撑,同时为在林分水平上的杨树病害生态调控技术的实施提供理论依据。
1 研究地区概况研究地点设在北京市大兴区西麻各庄大兴林场(116°22′42″E,39°42′10″N)。该场地处永定河东岸的平原地区,海拔45 m。暖温带季风气候,年平均气温11.5 ℃,年平均降水量556 mm,无霜期210天。土壤属于河滩土,有机质含量12~15 g·kg-1, 全氮含量0.8~1.0 g·kg-1,碱解氮含量60 ~75 mg·kg-1,速效磷含量15~30 mg·kg-1,速效钾含量50~100 mg·kg-1。
该地杨树人工林均为4年生107、108、DN113速生杨纯林,属黑杨派欧美杨杂交品种。其中107杨为相对高度抗病品种,108杨为中度抗病品种(王孟昌等,2008)。
2 研究方法 2.1 标准地设置与数据采集 2.1.1 不同品种不同指标标准地107杨、108杨和DN113杨的栽植密度为3 m×2 m (1 667株·hm-2),分别设立30 m×30 m标准地3块,共9块。所有标准地均为同龄林。
2.1.2 同一品种不同密度下不同指标标准地107杨株行距为3 m×2 m(1 667株·hm-2),4 m×2 m(1 250株·hm-2)和3 m×4 m (883株·hm-2),对每个处理设置30 m×30 m标准地3块,共9块。所有标准地均为同龄林。
2.1.3 调查时间与方法调查时间选在杨树病害的发病盛期。针对标准地内的活立木和枯死木逐株编号、定位,并调查记录胸径、树高、冠幅、树干距地面高度0.8~1.8 m范围内的病斑数量和病斑面积等。采用测高仪测定样地内每株树木的树高,用围尺测量胸径,测量树木南北方向的宽度来表示冠幅。
2.2 林木个体指标计量 2.2.1 大小比数大小比数是指林木个体指标大于参照树的相邻木占n株最近相邻木的株树比例,用以描述树种的生长优势程度(描述非均一性),用公式表示为(惠刚盈等,1999;郝云庆等,2006)

|
式中:Ui为第i株参照树的大小比数;Kij为一个离散性的变量,其值定义为当参照树i比相邻木j小时Kij=0,反之,Kij=1。大小比数量化了参照树与其相邻木的关系,Ui值越高,说明比参照树大的相邻木越多,参照树的优势度越大。Ui值越低,表明周围比参照树大的相邻木越少,参照树的优势度越小。大小比数取值有5种情况,即0(绝对劣势)、0.25(劣势)、0.5(中庸)、0.75(亚优势)和1(优势)。本文应用胸径、树高和冠幅来计测大小比数,即胸径大小比数、树高大小比数和冠幅大小比数,来描述树种的生长优势程度。
2.2.2 发病程度本文针对杨树干部溃疡病病害发生情况,通过测定树干、树干距地面高度0.8~1.8 m范围内的病斑数量、病斑面积等指标,记录当年新发的溃疡病病斑数量、估测病斑面积。
病害发生程度采用树干感病面积比(infected area ratio of trunk)描述,用来描述树木个体病害发生严重程度的指标,用单株树木树干高度0.8~1.8 m范围内病斑总面积占树干总表面积的百分比表示。数据处理时将树干感病面积比作转换,其中i表示第i株林木的树干感病面积比。
树干感病面积比(%)=0.8~1.8 m树干上病斑总面积/该段树干总表面积×100%。
2.3 数据分析测量数据经标准化后,用SPSS18.0软件的相应模块对108杨、107杨和DN113杨优势度指标(胸径大小比数、树高大小比数、冠幅大小比数、胸径、树高和冠幅)与溃疡病发病程度进行Person相关性分析和双尾显著性检验,采用方差分析比较不同品种和不同密度情况下,树木个体优势度间发病程度差异性,并进行多重比较(Tukey’s t-test, α=0.05)。
3 结果与分析 3.1 不同品种不同指标大小比数优势度与溃疡病发病程度的关系对测定的杨树的胸径、树高和冠幅的大小比数值与溃疡病发病情况进行相关性分析,结果表明:整体上,3个杨树品种的溃疡病发病程度与胸径、树高和冠幅的大小比数均呈极显著负相关(表 1),即个体优势程度越高,其发病程度越低。其中,108杨溃疡病发生情况与胸径大小比数相关性最高,相关系数为-0.672, 其次为DN113杨和107杨(rDN113= -0.595,r107=-0.560)。欧美杨DN113的发病情况与树高大小比数相关性高于108杨和107杨(rDN113=-0.620,r108=-0.471,r107=-0.406)。个体冠幅大小比数与溃疡病发病情况的相关系数大小:108杨>DN113杨>107杨。
|
|
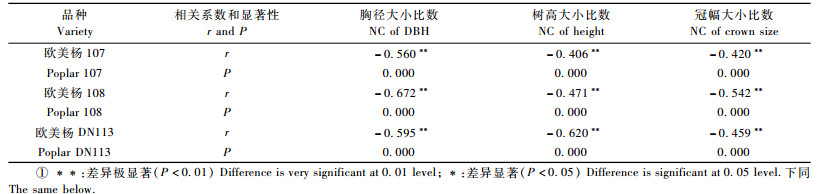
从图 1可知:在3 m×2 m株行距下,3种杨树品种的发病程度均与不同指标个体优势度呈显著的负相关性。108杨林木胸径和冠幅的个体优势度与溃疡病病情间的相关系数均大于DN113杨和107杨,树高的个体优势度与其的相关系数略低于DN113杨,但大于107杨。说明在该栽培模式下,108杨的感病情况受相邻木的影响较大,107杨最小,DN113杨介于其中,但这三者之间并无明显差异,表明在此栽培模式下,3个品种的感病情况均随着树木个体优势度的增大而降低。

|
图 1 不同品种杨树溃疡病发病程度与各大小比数间的关系 Fig.1 Relationship between neighborhood comparison(NC)and canker disease severity index of different poplar varieties |
方差分析表明:107杨、108杨和DN113杨溃疡病感病情况在不同指标个体优势度间均存在极显著差异(表 2)。在所有指标中,3个品种的发病程度在优势木和亚优势木(U=0.75,1)与绝对劣势木之间存在明显差异,劣势木的发病程度明显的高于优势木和绝对劣势木,说明处于优势状态下的树木比处于劣势状态下不容易发病;在绝对劣势木与中庸木间均有显著差异,而在中庸木和劣势木间,劣势木的发病程度普遍高于中庸木;在劣势木与绝对劣势木之间,虽然感病情况存在差异,但107杨的差异并不明显。因此,3个品种在林分中个体优势度处于劣势状态下,感病情况中庸木高,而且明显高于优势木或亚优势木。以树高和冠幅作为变量参数,108杨和DN113杨的发病程度在优势木、亚优势木和中庸木间无显著差异。
|
|
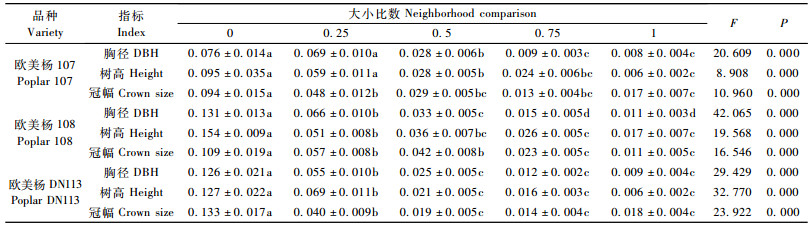
对不同密度下107杨的胸径、树高和冠幅的大小比数值与溃疡病发病情况进行相关性分析。结果表明:在3 m×2 m株行距下的发病程度与胸径大小比数相关性明显高于4 m×2 m株行距(r3×4=-0.553,r4×2=-0.362),而3 m×4 m株行距基本无相关性(r3×2=-0.129)。在3 m×2 m株行距下发病程度与树高和冠幅的大小比数均呈极显著负相关(表 3),另2个株行距下均无相关性,P>0.05。此外,在3 m×2 m和4 m×2 m的株行距下,各指标大小比数与发病程度的相关系数均大于不同指标的相关系数,而在3 m×4 m株行距下,与此正好相反。
|
|

由图 2可见:在株行距为3 m×2 m的林地中,杨树溃疡病病害与各指标的大小比数的相关性较为显著;溃疡病发病程度与胸径大小比、树高大小比和冠幅大小比均呈显著性负相关。但随着密度的降低,其相关性越来越弱,当株行距达到3 m×4 m时,杨树溃疡病病害的发生基本不受单株林木优势度的强弱影响。在林木空间结构中,不同指标的计测值比参照树大的最近相邻木个数越多,林分内个体间的竞争越激烈,树木生长势受到影响,感染杨树溃疡病的概率也会随之增大。

|
图 2 不同密度下欧美杨107杨树溃疡病发病程度与不同指标大小比数的关系 Fig.2 Relationship between neighborhood comparison(NC)and canker disease severity index of poplar 107 |
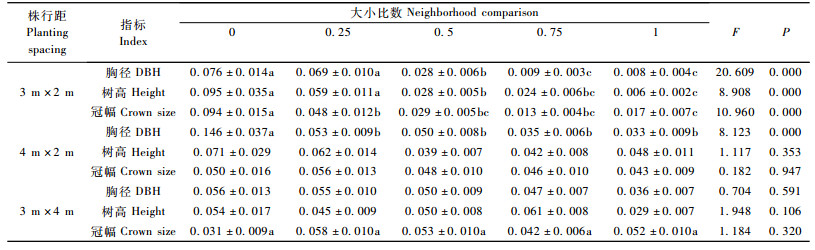
方差分析表明(表 4):在3 m×2 m株行距下,107杨溃疡病感病情况在不同指标个体优势度间存在极显著差异(胸径F=20.609,P<0.001;树高F=8.908,P<0.001;冠幅F=10.960,P<0.001);多重比较结果表明:以胸径和树高做变量参数,溃疡病发病程度在优势木和亚优势木间无显著性差异,劣势木和绝对劣势木间无显著差异、与中庸木、亚优势木和优势木间差异显著,同时亚优势木和优势木间无差异并均与中庸木差异显著。
在4 m×2 m栽培模式下,107杨溃疡病感病情况在胸径优势度间存在显著差异(胸径F=8.123,P<0.001);多重比较结果表明:除绝对劣势木外,其他几个优势度间无显著差异。树高和冠幅个体优势度间差异都不显著。
|
|
1) 在以往的研究中,大部分采用因子的变异系数和大小进行树木优势度的分化,这种单一的参数往往会因为缺乏必要的林分空间信息因子而造成分化的片面性,计算均值时易混淆。而大小比数着眼点放在各林木及其最近几株相邻木的关系上,能够准确地量化参照树与相邻木的大小关系,从而使在空间结构参数基础上人工重复复杂的林分结构成为可能。也就是说,单木的优势度取决于周围最近相邻木的大小,而同一级别的单木随相邻木的大小呈现不同的优势度。区域化的杨树人工林病害防治应当更加注重营林技术与病害防控技术相结合,从而达到病害的“生态控制”的最终目标(梁军等,2005)。在一定密度下,由于个体不同的优势度对发病程度起到关键作用,因此,在指导实施抚育管理措施中,以间伐个体优势度低(劣势木或绝对劣势木)的林木才能提高整体林分的生长潜力,同样可通过间伐个体优势度低的林木尤其是感病的优势度低的林木控制溃疡病病害。
2) 对特定的人工林而言,在林分水平上,密度因素有可能成为影响人工林树木个体树势的关键因素(Waring et al., 1982);而树势降低则可能进一步诱发病虫害的暴发。同时,人工林的种植由于品种单一,在景观水平上形成资源种类贫乏、分布过于集中等特征,因而使群体间的联合抗性作用降低,进而使得人工林的病虫害大规模暴发成为可能(Tahvanainen et al., 1972;Root,1973)。一般地,随着林分密度增加,密度效应会增强林分内个体间的竞争,林分中树木个体的树势有可能因个体间对空间、光照及水肥竞争加剧呈现整体降低的趋势(王静等,2011)。因此,密度对人工林林木群体生长发育过程起关键作用,林分密度减小,林木之间的距离增大,充足的阳光和空间保证了林分个体充分生长发育,树势得以提高,其抵抗力也随之增强,但在提高林木抵抗力的同时又无法最大限度地利用空间条件增加单产。因此,找到最佳的种植密度至关重要。
3) 区域化的杨树人工林病害防治应当更加注重营林技术与病害防控技术相结合,从而达到病害的“生态控制”的最终目标(梁军等,2005)。在一定密度下,由于个体不同的优势度对发病程度起到关键作用,因此,适时采取间伐措施,人为砍伐劣势木,有可能成为预防杨树人工林病害在林分水平上发生、流行与暴发的关键。
4) 描述林木生长情况有多个指标,逐项测定工作量较大。分析表明:林分胸径和树高的个体优势度与感病情况有较强的相关性,二者在反映生长优势方面是一致的。鉴于实测中准确的树高值很难测量,以后的研究中同种的树高大小比数可由胸径大小比数来代替,以减少工作量及降低测量难度,提高数据准确性。
| [] | 郝云庆, 王金锡, 王启和, 等. 2006. 柳杉人工林林分不同变量大小比数研究. 应用生态学报, 17(4): 751–754. |
| [] | 黄烈健, 苏晓华. 2003. 我国杨树溃疡病研究进展. 世界林业研究, 16(4): 49–53. |
| [] | 惠刚盈, Von GadowK, AlbertM. 1999. 一个新的林分空间结构参数——大小比数. 林业科学研究, 35(1): 1–6. |
| [] | 刚盈, Von Gadow K. 2003. 森林空间结构量化分析方法. 北京: 中国科学技术出版社. |
| [] | 惠刚盈, 胡艳波. 2001. 混交林树种空间隔离程度表达方式的研究. 林业科学研究, 14(1): 177–181. |
| [] | 雷相东, 唐守正. 2002. 林分结构多样性指标研究综述. 林业科学, 38(3): 146–141. |
| [] | 梁军, 张星耀. 2005. 森林有害生物生态控制. 林业科学, 41(4): 168–176. DOI:10.11707/j.1001-7488.20050429 |
| [] | 汤孟平, 唐守正, 雷相东. 2004. 两种混交度的比较分析. 林业资源管理, 4(8): 25–27. |
| [] | 王静, 孙志强, 梁军, 等. 2011. 中型景观尺度下杨树人工林林分特征对树干病害发生的影响——以河南省清丰县为例. 生态学报, 31(10): 2757–2767. |
| [] | 王孟昌, 梁军, 樊军锋, 等. 2008. 主要杨树生产品种对溃疡病田间抗性的调查. 西北林学院学报, 23(5): 122–123. |
| [] | 张群, 范少辉, 沈海龙, 等. 2003. 红松混交林中红松幼树生长环境的研究进展与展望. 林业科学研究, 16(2): 216–224. |
| [] | 张星耀, 骆有庆. 2003. 中国森林重大生物灾害. 北京, 中国林业出版社. |
| [] | Ferris R, Humphrey J W. 1999. A review of potential biodiversity indicators for application in British forests. Forestry, 72(4): 313–328. DOI:10.1093/forestry/72.4.313 |
| [] | Hu Y B, Hui G Y, Qi J Z, et al. 2003. Analysis of the spatial structure of natural Korean pine broad leaved forest. For Res, 16(5): 523–530. |
| [] | Larsson S. 1989. Stressful times of the plant stress: insect performance hypothesis. Oikos, 56(2): 277–283. DOI:10.2307/3565348 |
| [] | Mason W L, Quine C P. 1995. Silvicultural possibilities for increasing structural diversity in British spruce forests the case of Kielder forest. Forest Ecology and Management, 79(1): 13–28. |
| [] | Pommerening A. 2002. Approaches to quantifying forest structures. Forestry, 75(3): 305–324. DOI:10.1093/forestry/75.3.305 |
| [] | Root R B. 1973. Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecological Monographs, 43(1): 95–124. DOI:10.2307/1942161 |
| [] | Tahvanainen J O, Root R B. 1972. The influence of vegetational diversity on the population ecology of a specialized herbivore, Phyllotreta cruciferae (Coleoptera: Chrysomeloidae). Oecologia, 10(4): 321–346. DOI:10.1007/BF00345736 |
| [] | Waring R H, Schroeder P E, Oren R. 1982. Application of the pipe model theory to predict canopy leaf area. Journal of Forest Research, 12: 556–560. |