 2012, Vol. 48
2012, Vol. 48文章信息
- 王玉平, 陶建平, 刘晋仙, 何泽
- Wang Yuping, Tao Jianping, Liu Jinxian, He Ze
- 不同光环境下6种常绿阔叶林树种苗期的叶片功能性状
- Response of Leaf Functional Traits to Different Light Regimes in an Evergreen Broad-Leaved Forest in the Jinyun Mountain
- 林业科学, 2012, 48(11): 23-29.
- Scientia Silvae Sinicae, 2012, 48(11): 23-29.
-
文章历史
- 收稿日期:2011-12-06
- 修回日期:2012-09-29
-
作者相关文章



植物对环境变化的响应及适应策略一直以来都是生态学研究的热点问题之一。植物功能性状(plant functional traits)是指能够响应生存环境的变化,并对生态系统具有一定影响的植物属性(Díaz et al., 2001),它与环境的联系是气候、干扰和生物条件筛选的结果(Díaz et al., 1998)。叶片是绝大多数维管植物能量生产和物质积累的中介,同时植物对环境变化的绝大多数响应也表现在叶级性状(leaf-level traits)(Valladares et al., 2000)。在各种植物功能性状中,叶功能性状(leaf functional traits)能够直接反映植物适应环境变化所形成的生存对策,与植株生物量和植物对资源的获得、利用及利用效率的关系密切(孟婷婷等, 2007),同时,叶片性状容易收集,便于进行大尺度的比较研究(Hodgson et al., 1999)。因此,研究者已在叶片功能性状方面进行了大量的研究(Wright et al., 2004a; 2004b; He et al., 2006),但多集中在全球不同气候带或不同群落类型等大(或较大)尺度上(Wright et al., 2004a; Suding et al., 2008; 宝乐等, 2009)。在大尺度下,环境对植物功能性状,特别是叶片功能性状的影响往往不大(Wright et al., 2004b; 2005; He et al., 2006),且大尺度下叶片性状间的关系不能完全在区域等小尺度上受到重现(Wright et al., 2004b)。因此,在大尺度研究的背景下,小尺度(区域性)控制试验的开展在提供叶片功能性状对环境变化响应的细节方面具有重要意义。目前这方面的研究还相对较少,且主要集中在人为遮光和养分控制等方面(肖春旺等, 1999; Valladares et al., 2000; 万宏伟等, 2008)。
本文通过试验生态学的方法,对6种缙云山常绿阔叶林常见树种在林下、人工林窗及林缘旷地中树种苗期在不同光环境下,叶片性状变化规律及性状间关系的变化;不同演替阶段物种的叶性状对光环境的响应是否存在差异进行探讨。
1 材料与方法 1.1 研究地概况研究地位于西南大学三峡库区生态与环境教育部重点实验室试验基地内,海拔249 m,气候属典型的亚热带季风气候,年均气温13.6 ℃,年均降水量1 143 mm,土壤为酸性黄壤,距离缙云山保护区直线距离约2 km,气候和土壤条件相近。
1.2 试验材料试验材料为薄果猴欢喜(Sloanea leptocarpa)、喜树(Camptotheca acuminata)、香樟(Cinnamomum camphora)、黑壳楠(Lindera megaphylla)、薯豆(Elaeocarpus japonicus)、灯台树(Bothrocaryum controversum)。灯台树、喜树和香樟为演替前期种,喜光;薄果猴欢喜、薯豆和黑壳楠为演替后期种,具有耐荫特性(郭全邦等, 1999)。
1.3 材料处理方法2005年3月(2006年补栽了部分幼苗),从缙云山自然保护区采集长势一致、生长良好的6种试验材料当年生幼苗各30株,种植于直径25 mm,深20 cm的营养袋中,以缙云山林下带腐殖质的土壤为前期栽培基质。在林窗中恢复生长2周后,将幼苗随机分成3组,分别种植于试验基地的常绿阔叶林林缘旷地、常绿阔叶林人工林窗和林下3种自然光环境下。3种环境的相对光照强度分别为林缘旷地100%、人工林窗18%~23%、林下3%~7%,其有效光辐射的日变化规律参见汤爱仪等(2008)报道。对材料进行常规管理,同时对林窗冠层进行不定期修剪,保持林窗形态。
1.4 采样处理及分析方法2010年7月,时间选择在18:00左右,每种随机选取3~5株个体,采集冠层当年生枝条上完整的成熟叶片,分别装入信封,迅速带回实验室测量相关指标。由于自然状态下的长时间处理,部分材料在林窗(喜树和猴欢喜)和林下(香樟和喜树)环境中死亡或数量不足,故林窗和林下环境中只选择了其余4种试验树种。
1.4.1 比叶面积和叶片干物质含量的测定用根系扫描仪(EPSON Expression 10000XL, USA)扫描叶片,用WinRhizoRegV2004a分析叶片投影面积,然后放入60 ℃的烘箱中,烘干至恒质量(72h),称量干质量,计算比叶面积(specific leaf area, SLA, cm2·g-1,叶面积/叶干质量)。叶片干物质含量(leaf dry matter content, LDMC)是指单位饱和鲜质量的叶片所含干物质的多少(Cornelissen et al., 2003)。将叶片放入黑暗环境的清水中浸泡,12h后取出叶片并用吸水纸将叶片表面的水分吸干,同时进一步将叶片杂质擦拭干净,称量饱和鲜质量,然后在60 ℃下烘干至恒质量(72h),称量干质量, 计算LDMC (叶干质量/饱和鲜质量,g·g-1)。
1.4.2 叶片N, P含量的测定将烘干的叶片用混合球磨仪(MM200, Retsch, Germany)磨碎混匀,磨碎前去除叶柄和明显叶脉。用浓H2SO4-H2O2消煮材料,然后采用凯氏定氮仪(B-324, Buchi, Switzerland)测定干样全N含量,钼锑抗比色法测定干样全磷含量(鲍士旦, 2000)。N,P测定结果以单位质量样品的养分含量(mg·g-1)表示。
1.5 数据处理与统计分析对叶片性状指标数据进行光环境和演替阶段的双因素方差分析,对差异显著(P < 0.05)的指标作Duncan多重比较;不同演替阶段物种间的比较用独立样本t检验;性状间的相关性采用Pearson双侧检验。应用软件SMATR (Falster et al., 2006)对不同光环境下叶功能性状之间的关联进行了Ⅱ型回归分析(Model Ⅱ Regression);对原始数据进行了底数为10的对数变换以保证正态分布。
2 结果与分析 2.1 树种叶片功能性状对不同光环境的响应除叶片磷含量(leaf phosphorus content, LPC)外,比叶面积(SLA)、叶片干物质含量(LDMC)、叶片氮含量(leaf nitrogen content, LNC)及氮磷比(N:P)在不同光环境下具有极显著差异(P < 0.01);不同演替阶段物种间叶片性状均存在极显著差异(P < 0.01)。另外,2种因素的交互作用只对比叶面积的影响达到极显著水平(P < 0.01),对其他叶片性状影响不显著(表 1)。
|
|
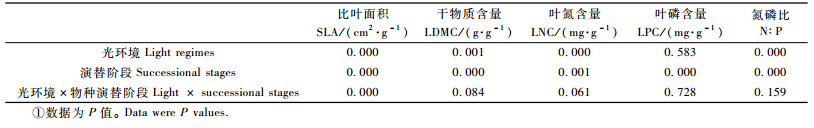
SLA随着光照的减弱,总体呈先增加后降低的趋势。林下和林窗环境下SLA极显著地高于旷地环境下植物的SLA(P < 0.01),林窗与林下之间无显著差异(P>0.05)(表 2)。LDMC在3种环境下的变化趋势为先降低后增加;旷地和林窗与林下LDMC无显著差异,林窗中LDMC极显著低于旷地环境(P < 0.01)。
|
|
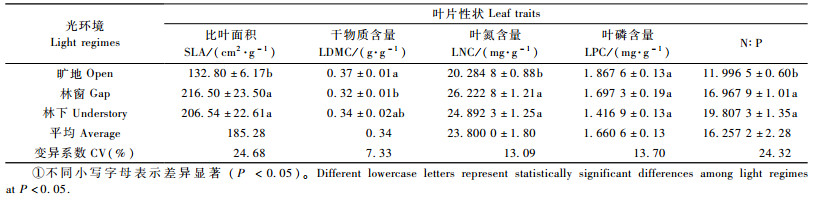
3种光环境下,LNC变化幅度为20.3~26.2 mg·g-1,林窗和林下的LNC显著高于旷地(P < 0.05)。LPC变化幅度为1.42~1.87 mg·g-1,且随着光照的减弱依次降低,但三者间无显著差异(P=0.117)。N:P比变化幅度较大(12.0~19.8)。随着光照的减弱,N:P比逐渐增大,且旷地环境下N:P比显著低于林窗和林下环境(P < 0.05)。
2.1.3 不同演替阶段物种在不同光环境下的比叶面积和叶干物质含量演替前期种的SLA在3种光环境下均高于演替后期种,其中在林窗和林下,差异达到显著水平(P < 0.05);而LDMC则相反,3种环境下均为演替后期种显著高于演替前期种(P < 0.05)。随着光照的减弱,演替前期种SLA逐步增大,LDMC减小,且林下和旷地间差异显著(P < 0.05);演替后期种随着光照的减弱,SLA先增加后减小,LDMC变化趋势相反(先减少后增加),且两者均在旷地环境下与林窗和林下差异显著(P < 0.05)。
2.1.4 不同演替阶段物种在不同光环境下植物叶片N,P含量及N:P特征不同演替阶段物种的LNC在旷地(P=0.121)和林窗(P=0.066)环境下无显著差异,而在林下演替前期种显著高于演替后期种(P < 0.05)(图 2A);在3种光环境下,演替前期种的LPC均显著高于演替后期种(图 2B);N:P比为演替后期种均显著高于演替前期种(图 2C)。随着光照的减弱,演替前期种的LNC逐渐增大,而演替后期种在林窗环境下最大,但均表现为在旷地环境下显著低于林窗和林下;LPC表现为演替后期种随着光照的减弱呈下降趋势,且旷地和林下间差异显著(P < 0.05),而演替前期种在3种光环境下组间差异不显著;随着光照的减弱,N:P比逐渐增大,但演替前期种在林下和旷地间差异显著,演替后期种林下和林窗均显著高于旷地。

|
图 1 不同演替阶段物种在3种光环境下比叶面积和叶干物质含量 Fig.1 SLA and LDMC of species at different successional stages under different light regimes 小写字母表示不同演替阶段物种性状在同一光环境下的差异;大写字母表示同一演替阶段物种性状在不同光照环境下的差异。 Lowercase letters represent the differences in leaf traits between successional stages under same light regime; capital letters represent the differences in leaf traits among light regimes for the same successional stage species.下同.The same below. |

|
图 2 不同演替阶段物种在不同光环境下植物叶片N,P含量及N:P比 Fig.2 LNC, LPC and N:P ratio of species at different succession stages under different light regimes |
在3种光环境下,LNC与LPC均呈显著的正相关关系(图 3A),标准主轴(SMA)共同斜率为1.76。不同光环境下,LNC-LPC关系沿共同斜率无显著变异,而林窗和林下的Y轴截距显著小于旷地(表 3),即在给定LNC情况下,随着光照的减弱,LPC降低。

|
图 3 不同光环境下叶片功能性状间的关系(Ⅱ型回归) Fig.3 Relationships between leaf functional traits across different light regimes (Model Ⅱ regression) 数据进行了10为底的对数转换。 All data were lg-transformed before analysis. |
|
|
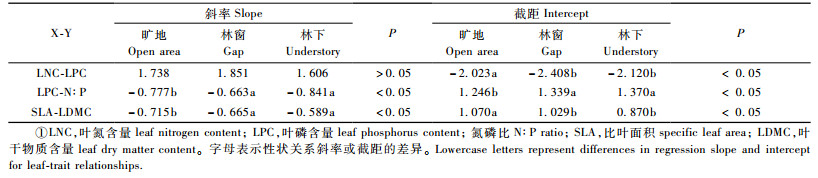
N:P比与LPC呈显著的负相关关系,SMA共同斜率为-0.74。不同光环境下,N:P-LPC关系沿共同斜率发生了显著的变异,林窗和林下的Y轴截距显著大于旷地,即随光照降低,物种具有更小的LPC和更大的N:P值(图 3B,表 3)。
SLA与LDMC呈显著的负相关关系,SMA共同斜率为-0.64。不同光环境下,SLA-LDMC关系沿共同斜率发生了显著变异,林窗和林下的Y轴截距显著低于旷地,表明随光照的降低,物种具有更大的SLA和较小的LDMC(图 3C,表 3)。
2.2.2 比叶面积、叶干物质含量与叶片养分含量的关系LNC和LPC与LDMC均呈显著的负相关关系(R2=0.378,P < 0.05;R2=0.603,P < 0.01),LPC与LDMC相关性更显著(图 4A, B)。SLA与LNC呈显著的正相关关系(R2=0.469,P < 0.01)(图 4C),与LPC关系不显著。

|
图 4 比叶面积、叶干物质含量与叶片养分含量的关系 Fig.4 Relationships between SLA, LDMC and leaf nutrient contents |
植物的叶片性状反映了植物对环境变化的不同响应策略(刘金环等, 2006)。比叶面积和叶干物质含量都能在一定程度上代表植物对环境资源的利用方式和能力,与环境适应对策关系密切(Niinemets, 2001),被认为是环境资源利用轴(resource use axis)上的一组最佳变量(刘金环等, 2006)。比叶面积能够反映植物对资源获取和利用的权衡关系(Wilson et al., 1999; Long et al., 2011)。研究表明:在荫蔽的林下植物有较大的比叶面积(Valladares et al., 2000; Cornelissen et al., 2003)。叶干物质含量反映的是植物对养分的保持能力(祁建等, 2008),叶干物质含量较低的植物叶片具有较强的生产能力(Cornelissen et al., 2003)。本研究中,林窗和林下环境中的比叶面积显著高于林缘旷地,叶干物质含量在3种光环境下变化趋势与比叶面积相反(表 2),表明植物叶片在荫蔽的环境下,一方面通过增大受光面积来捕捉更多的光资源;另一方面,通过增强叶片的生产能力和养分保守能力,高效地利用光资源。
氮和磷是陆地生态系统植物生长的主要限制因子(任书杰等, 2007)。叶氮是光合酶(特别是Rubisco)和叶绿素的主要成分,与磷的固定能力呈正相关(赵平等, 1998; Wright et al., 2004a);叶磷与植物的生长速率密切相关(曾德慧等, 2005)。在光资源不足的林下,植物叶片通过增加单位干质量的氮含量来提高净光合速率(赵平等, 1998),同时,植物体生长减缓,叶片内限制磷含量的核糖体(特别是rRNA)对磷的需求降低(程滨等, 2010),因此叶磷含量随之降低,同时N:P比升高(表 2)。在本研究中,叶磷含量随光照减弱而降低,但差异不显著,这可能与弱光光环境下成熟叶片生命活动周期延长有关(Niinemets, 2001)。
叶片的N:P比值可以作为判断植物生长是否受到养分限制(Güsewell, 2004)。Koerselman等(1996)研究认为:当N:P比小于14时,表示植物生产力受N的限制;N:P比大于16时,表示植物生产力受P的限制;介于中间则被认为受两者共同限制或2种元素都不缺少(Tessier et al., 2003)。本研究中,旷地环境下植物叶片N:P比小于14,在林窗和林下环境下,分别为17和20,而本研究中土壤养分环境基本一致,由此说明,光照显著影响了N:P比对环境养分限制的指示作用。小尺度范围内,在利用N:P临界比判断环境或系统养分限制状况的研究中,应考虑光环境,因为植物生物量的生产除N,P元素限制外,还将受到光照和其他气候因子的影响(Duren et al., 2000)。
3.2 不同演替阶段的物种对光环境的响应本研究中,在3种光环境下,演替后期种的叶干物质含量和N:P比均显著高于演替前期种,叶磷含量相反,这体现了2种演替阶段的物种本身所具有的差异:演替前期种生长快速,后期种具有较强的养分保持能力和耐阴能力。在光资源丰富的旷地,演替前期和后期种的比叶面积和叶氮含量均无显著差异。随着光照的减弱,不同演替阶段物种的比叶面积在林窗和林下出现显著差异,叶氮含量在林下出现显著的差异,表明在光资源足够的环境下比叶面积和叶氮含量与物种演替阶段关系不大;而当光资源减少(林窗)或成为限制因子时(林下),不同演替阶段物种的叶性状出现差异性响应,说明了比叶面积和叶氮含量两性状对光资源有较好的指示作用。
不同演替阶段物种对光环境变化的响应也不一致。演替前期物种的叶磷含量随光照的减弱而无显著差异,而演替后期种则逐渐降低,且林下与旷地差异显著,表明演替前期种的快速生长策略,同时跟演替前期种在3种光环境下都具有较大的比叶面积和叶N含量有关。
3.3 植物叶片功能性状间的关系与功能性状和气象因子间的关系相比,功能性状间的关系更加紧密(Wright et al., 2005)。在全球尺度上,比叶面积、叶寿命、叶碳含量、叶磷含量及单位物质的光合能力等互相具有密切的相关性,且具有全球普遍性(Reich et al., 1997; Wright et al., 2004a)。大量研究表明:比叶面积与叶干物质含量呈负相关关系(Wilson et al., 1999; Wright et al., 2004b);叶氮含量与叶磷含量呈正相关关系;N:P比与叶P含量呈负相关关系而与叶N含量无显著相关性(高三平等, 2007),本研究结果与这些报道一致(图 3)。同时,叶片养分含量与植物资源利用或植物适应环境的策略也存在一定的相关关系(冯秋红等, 2010)。叶氮含量与比叶面积呈正相关,与叶干物质含量呈负相关关系(图 4),表明随着比叶面积的增大,叶干物质含量减小,同时分配到单位质量光合器官的氮也随之增加,以维持较高的光合能力。
性状间的关系还受环境因子的影响(冯秋红等, 2010)。本研究中,随着光照的减弱,SLA-LDMC的关系逐渐增强,且沿着共同斜率和Y轴截距均发生了显著的变异,趋于更大的比叶面积和较小的叶干物质含量(图 3C);LNC-LPC的关系在林窗下最显著,在林窗和林下环境下Y轴截距显著小于旷地(图 3A);LPC-N:P关系随着光照的减弱向高氮磷比、低叶磷含量方向移动(图 3B)。表明植物性状的权衡关系沿着光资源梯度发生了相应的动态变化以适应不同光照环境(冯秋红等, 2008),即环境因子通过改变植物性状来影响植物的同时,还通过控制和影响植物性状间的关系来影响植物的适应策略。
| [] | 宝乐, 刘艳红. 2009. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 29(7): 3692–3703. |
| [] | 鲍士旦. 2000. 土壤农化分析. 3版. 北京: 中国农业出版社. |
| [] | 程滨, 赵永军, 张文广, 等. 2010. 生态化学计量学研究进展. 生态学报, 30(6): 1628–1637. |
| [] | 冯秋红, 史作民, 董莉莉. 2008. 植物功能性状对环境的响应及其应用. 林业科学, 44(4): 125–131. DOI:10.11707/j.1001-7488.20080423 |
| [] | 冯秋红, 史作民, 董莉莉, 等. 2010. 南北样带温带区栎属树种功能性状间的关系及其对气象因子的响应. 植物生态学报, 34(6): 619–627. |
| [] | 郭全邦, 刘玉成, 李旭光. 1999. 缙云山森林次生演替序列群落的物种多样性动态. 应用生态学报, 10(5): 521–524. |
| [] | 高三平, 李俊祥, 徐明策, 等. 2007. 天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征. 生态学报, 27(3): 948–952. |
| [] | 刘金环, 曾德慧, LeeD K. 2006. 科尔沁沙地东南部地区主要植物叶片性状及其相互关系. 生态学杂志, 25(8): 921–925. |
| [] | 孟婷婷, 健倪, 王国宏. 2007. 植物功能性状与环境和生态系统功能. 植物生态学报, 31(1): 150–165. |
| [] | 祁建, 马克明, 张育新. 2008. 北京东灵山不同坡位辽东栎(Quercus liaotungensis)叶属性的比较. 生态学报, 28(1): 122–128. |
| [] | 任书杰, 于贵瑞, 陶波, 等. 2007. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 28(12): 2665–2673. DOI:10.3321/j.issn:0250-3301.2007.12.001 |
| [] | 汤爱仪, 陶建平, 刘欣, 等. 2008. 常绿阔叶林两种树种幼苗对不同光环境的适应——关于生长特征和光合色素含量的研究. 西南大学学报:自然科学版, 30(1): 109–113. |
| [] | 万宏伟, 杨阳, 白世勤, 等. 2008. 羊草草原群落6种植物叶片功能特性对氮素添加的响应. 植物生态学报, 32(3): 611–621. |
| [] | 肖春旺, 刘玉成. 1999. 不同光环境下四川大头茶幼苗的生态适应. 生态学报, 19(3): 422–426. |
| [] | 阎恩荣, 王希华, 周武. 2008. 天童常绿阔叶林演替系列植物群落的N:P化学计量特征. 植物生态学报, 32(1): 13–22. |
| [] | 曾德慧, 陈广生. 2005. 生态化学计量学:复杂生命系统奥秘的探索. 植物生态学报, 29(6): 1007–1019. |
| [] | 赵平, 孙谷畴. 1998. 植物氮素营养的生理生态学研究. 生态科学, 17(2): 37–41. |
| [] | Cornelissen J H C, Lavorel S, Garnier E, et al. 2003. A hand book of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 51(4): 335–380. DOI:10.1071/BT02124 |
| [] | Díaz S, Cabido M. 2001. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology and Evolution, 16(11): 646–655. DOI:10.1016/S0169-5347(01)02283-2 |
| [] | Díaz S, Cabido M, Casanoves F. 1998. Plant functional traits and environmental filters at a regional scale. Journal of Vegetation Science, 9(1): 113–122. DOI:10.2307/3237229 |
| [] | Falster D S, Warton D I, Wright I J. 2006. User's guide to SMATR: standardised major axis tests & routines, Version2.0, Copyright. http://www.bio.mq.edu.au/ecology/SMATR20. |
| [] | Güsewell S. 2004. N:P ratios in terrestrial plants: variation and functional significance. New Phytologist, 164(2): 243–266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [] | He J S, Wang Z, Wang X, et al. 2006. A test of the generality of leaf trait relationships on the Tibetan Plateau. New Phytologist, 170(4): 835–848. DOI:10.1111/nph.2006.170.issue-4 |
| [] | Hodgson J G, Wilson P J, Hunt R, et al. 1999. Allocating C-S-R plant functional types: a soft approach to a hard problem. Oikos, 85(2): 282–294. DOI:10.2307/3546494 |
| [] | Koerselman W, Meuleman A F M. 1996. The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 33(6): 1441–1450. DOI:10.2307/2404783 |
| [] | Long W X, Zang R G, Schamp B S, et al. 2011. Within-and among-species variation in specific leaf area drive community assembly in a tropical cloud forest. Oecologia, 167(4): 1103–1113. DOI:10.1007/s00442-011-2050-9 |
| [] | Niinemets V. 2001. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology, 82(2): 453–469. DOI:10.1890/0012-9658(2001)082[0453:GSCCOL]2.0.CO;2 |
| [] | Wright l J, Reich P B, Westoby M, et al. 2004a. The worldwide leaf economics spectrum. Nature, 428(6985): 821–827. DOI:10.1038/nature02403 |
| [] | Wright I J, Groom P K, Lamont B B, et al. 2004b. Leaf trait relationships in Australian plant species. Functional Plant Biology, 31(5): 551–558. DOI:10.1071/FP03212 |
| [] | Wright I J, Reich P B, Cornelissen J H C, et al. 2005. Modulation of leaf economic traits and trait relationships by climate. Global Ecology and Biogeography, 14(5): 411–421. DOI:10.1111/geb.2005.14.issue-5 |