 2012, Vol. 48
2012, Vol. 48文章信息
- 赵中华, 惠刚盈, 胡艳波, 李远发
- Zhao Zhonghua, Hui Gangying, Hu Yanbo, Li Yuanfa
- 树种多样性计算方法的比较
- Comparison of Tree Species Diversity Calculated
- 林业科学, 2012, 48(11): 1-8.
- Scientia Silvae Sinicae, 2012, 48(11): 1-8.
-
文章历史
- 收稿日期:2012-01-17
- 修回日期:2012-04-09
-
作者相关文章



生物多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的综合(Pielou,1975;马克平,1993),是生态系统生产力的核心(赵士洞等,1997),目前已成为全球生态学研究的热点问题。生物多样性的测度始于20世纪,Fisher等(1943)提出α多样性指数,用群落中的物种数目和每一个物种的个体数目来表达物种多样性;Simpson(1949)提出多样性的概率度量方法;Margalef(1958)首次把Shannon-Wiener的信息论指数用于生态学群落物种多样性的研究;Whittaker(1977)将多样性指数划分为4个空间尺度:α多样性、β多样性、γ多样性和δ多样性,其中,α多样性指数可分为4类,即物种丰富度指数、物种相对多度模型、物种丰富度与相对多度综合形成的指数以及物种均匀度指数(马克平,1994)。随着物种多样性定量研究的不断深入,目前至少出现30多个不同的生物多样性指数模型,许多文献对生物多样性指数的性质、相关性进行了讨论与评述(Magurran,1988;马克平等,1994;岳天祥,1999)。近年来,又出现了描述多样性的新方法,如Ganeshalah等(2000)在Shannon-Wiener的基础上提出雪崩指数(avalanche index);我国学者王永繁等(2002)提出新的α多样性测度指标DIV多样性指数;惠刚盈等(2011)提出基于相邻木空间关系的树种多样性测度方法——树种空间多样性指数(TSS)。
物种多样性是生物多样性在物种水平上的表现形式,是反映群落组织化水平,继而通过结构与功能的关系间接反映群落功能特征的重要指标(赵士洞等,1997)。对物种多样性的研究旨在揭示生物群落客观存在的物种和结构的多样性,并通过结构和功能间相关关系的分析进而揭示和认识生物群落的功能多样性。本文选择最新的树种多样性TSS指数和应用最为广泛的α多样性指数,以模拟数据、甘肃小陇山松栎混交林林区固定监测样地数据和抽样调查数据为例,比较分析各多样性测度指标的优缺点,以期为森林经营者提供参考。
1 研究数据 1.1 模拟数据为客观评价各物种多样性测度公式的不同,特设计如下3组模拟林分(图 1)。第Ⅰ和Ⅱ为2个树种等分空间的情景,第Ⅲ组为3个树种等分空间的情景。各组株数为,Ⅰ为64株,Ⅱ和Ⅲ的总计算株数均为108株。各组中又分a,b,c,d 4种情况,其中,a表示单株混交,b双株混交,c单行混交,d双行混交。在同一类型中,物种空间隔离程度的高低依次为a>b>c>d。

|
图 1 模拟林分 Fig.1 Fictitious forest |
实测数据来自甘肃省小陇山林区。小陇山林区位于甘肃省东南部,地处秦岭西端,我国华中、华北、喜马拉雅、蒙新四大自然植被区系的交汇处,地理坐标为33°30′—34°49′ N,104°22′—106°43′ E,属暖温带向北亚热带过渡地带,兼有我国南北气候特点,大多数地域属暖温湿润-中温半湿润大陆性季风气候类型。特殊的地理位置与环境条件、生物的地理成分与区系成分复杂多样,使小陇山林区成为甘肃省生物种质资源最丰富的地区之一。小陇山林区海拔2 200 m以下主要分布以锐齿栎(Quercus aliena var. acuteserrata)和辽东栎(Quercus wutaishanica)为主的天然林,在栎林带内分布华山松(Pinus armandi)、油松(Pinus tabulaeformis)、山杨(Populus davidiana)、漆树(Toxicodendron verniciflum)、冬瓜杨(Populus purdomii)、千金榆(Carpinus cordata)、甘肃山楂(Crataegus kansuensis)和刺楸(Kalopanax septemlobus)等乔木树种,灌木有美丽胡枝子(Lespedeza formosa)、中华绣线菊(Spiraea chinensis)、胡颓子(Elaeagnus pungens)、华北绣线菊(Spiraea fritschiana)、连翘(Forsythia suspensa)、卫矛(Euonymus alatus)和绒毛胡枝子(Lespedeza tomentosa)等。在小陇山林区百花林场王安沟设立1块3 600 m2的锐齿栎天然林样地,在小阳沟营林区设立2块4 900 m2的锐齿栎天然林样地,在李子林场采用无样地抽样技术设立9块不同林分类型临时样地。
在小陇山林区不同林分类型中设立的3块固定监测样地林分类型均为锐齿栎天然林,其中,王安沟样地(样地1)地理位置较偏僻,周围2 km范围内没有取材道,没有发现人为采伐的痕迹。样地内存在枯立木35株,胸径在20 cm以上的为11株,20 cm以下的为24株;枯倒木162株,20 cm以上的93株,20 cm以下的69株,可认为是近原始林。小阳沟营林区的2块固定样地(样地2和样地3)为锐齿栎天然次生林。样地4为1969年皆伐后天然更新的锐齿栎天然林,样地5为锐齿栎天然林经2次择伐后形成的林分,择伐时间分别为1980年和1995年。样地6~12为小陇山林区不同类型改造模式林分,其中,样地6,7分别为灌木林带状改造的华山松和油松模式,样地8,9为锐齿栎天然林皆伐后改造的华山松和油松模式,样地10~12为灌木林皆伐后改造的华山松、油松和日本落叶松(Larix kaempferi)模式。表 1为各调查林分的特征。由表 1可以看出,人为影响较少的锐齿栎天然林(样地1)的林分密度较大,为1 336株·hm-2,树种数则达到44种;锐齿栎天然次生林(样地2,3)和皆伐后天然更新的锐齿栎天然林(样地4)的树种数相对较多,树种数分别达到32种和29种;锐齿栎择伐林(样地5)的树种数为20种;天然林人工改造类型(样地6~12)的树种数相对较少,林分以改造树种为主,其中,天然灌林带状改造类型(样地6,7)的树种数较锐齿栎天然林全面改造人工林的树种多,天然灌木全面改造类型(样地9~11)的树种最少,为7种。
|
|

对3块固定监测样地运用TOPCON全站仪每木定位并全面调查;对临时样地采用无样地抽样调查,调查内容包括郁闭度、断面积、坡度、林分平均高、树种、直径及其结构参数(包括角尺度、混交度和大小比数)。树种、直径及空间结构参数调查采用点抽样的方法,即从1个随机点开始,每隔一定距离(以调查的参照树的最近4株相邻木不重复为原则)设立1个抽样点,以激光测角器和角规作为辅助设备,调查距抽样点最近4株胸径大于5 cm树木的树种、胸径级及角尺度、大小比数和混交度等空间结构参数,同时调查参照树与相邻树构成的结构单元的成层性和树种数,并用角规绕测林分断面积。每个样地抽样点为49个,每个抽样点涉及4株参照树,每个参照树涉及4株相邻木,角规绕测点5个以上。计算机模拟验证、在东北红松(Pinus koraiensis)阔叶林样地验证以及厄瓜多尔天然林样地验证均表明:采用49个点作为天然林林木空间分布格局调查的最小样本量是合理的(徐海等,2007;惠刚盈等,2007);对于人工林而言,由于组成结构简单,抽样点达到20个就能满足调查要求。
2.2 数据分析方法对调查林分的乔木层树种多样性应用目前最为广泛的生物多样性测度指数(马克平,1994)进行计算:
物种丰富度(S):
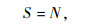
|
(1) |
Shannon-Wiener指数(H′)(Shannon et al., 1963):
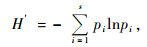
|
(2) |
Simpson指数(D)(Simpson, 1949):
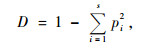
|
(3) |
MeIntosh指数(Dm)(McIntosh, 1967):

|
(4) |
Fisher指数(α)(Fisher et al., 1949):
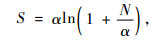
|
(5) |
式中:N代表树种的个体数;pi表示第i个种的个体占所有个体数的比例。
惠刚盈等(2011)提出了基于相邻木空间关系的树种空间多样性指数(TSS),该指数建立在结构单元参照树与其邻体的物种关系的基础上,其表达方式如下:
树种空间多样性指数(TSS):
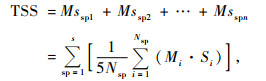
|
(6) |
式中:Mi为结构单元中的树种混交度,其计算公式为:
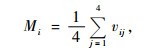
|
(7) |
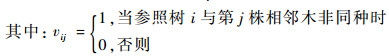
|
Mssp为各树种的平均空间状态,计算公式为:
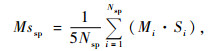
|
(8) |
式中:Nsp为树种sp个体数,Si为结构单元中的树种数,i为以树种sp为参照树的结构单元数。由(6)式可知,当群落由N个个体、N个物种组成,也就是说群落中每个物种个体只有1株时,该群落的物种多样性达最大值,等于物种丰富度(S)即TSS=S;当群落仅由1个物种的N个个体组成时,该群落的物种多样性达最小值即TSS=0。TSS与样地大小无关。(6)式是群落中所有物种的平均空间状态的集合,是群落中物种多样性的空间测度,故称其为物种空间多样性指数。
3 结果与分析 3.1 模拟林分的树种多样性表 2列举了6种多样性指数对模拟林分数据的计算结果。可以看出,MeIntosh指数(Dm)和Fisher指数(α)计算的物种多样性Ⅰ比Ⅱ大,与通常人们对多样性的理解相悖,而物种丰富度指数(S)、Shannon-Wiener指数(H′)和Simpson指数(D)以及物种空间多样性指数(TSS)计算的Ⅰ和Ⅱ的多样性与通常的理解相一致;6种多样性指数都能够表达出Ⅲ的多样性比Ⅰ,Ⅱ高这样一个事实,但在6种多样性指数中只有物种空间多样性指数(TSS)能够正确表达出在相同物种数量的群落中,由于物种的空间隔离程度不同而表现出不同的空间多样性,即在模拟林分中,仅有物种空间多样性指数表达出物种隔离程度a>b>c>d的高低顺序。
|
|
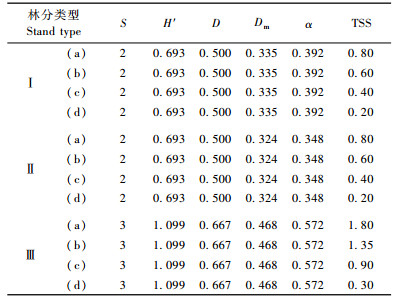
对各调查林分乔木层树种选择5种常用的多样性指数和惠刚盈等(2011)提出的基于相邻木关系的树种空间多样性指数进行分析(表 3)。
|
|
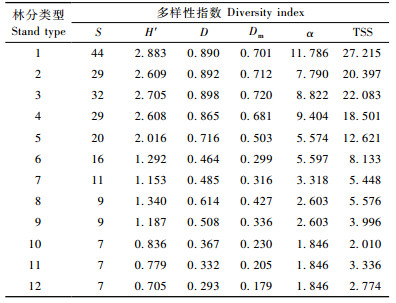
由表 3可见,Shannon-Wiener指数(H′)、Simpson指数(D)、MeIntosh指数(Dm)、Fisher指数(α)以及物种空间多样性指数(TSS)均随物种丰富度(S)的增加而增加,但增加的方式不尽相同,其中,H′,D和Dm出现了树种数为9(林分类型8和9)反而比树种数16(林分类型6)或11(林分类型7)的物种多样性高这样令人难以置信的现象;α在相同的调查株数和相同的树种数量时表现为相同的物种多样性,而没有像其他指数那样反映由于均匀度或多度的不同而出现物种多样性的不同,可见,α表达的是物种丰富度而并非物种多样性;TSS对群落物种多样性的计算结果总体上呈现随着树种丰富度的增加而增加,但也出现个别树种数为9(林分类型8)时树种多样性大于树种数为11(林分类型7)时的情况。此外,在树种均为7种时,出现了H′和TSS的计算结果相悖的结果,如表 3中林分类型(10)和(11)的情况。由Shannon-Wiener指数(H′)、Simpson指数(D)和MeIntosh指数(Dm)的计算公式可以看出,这几个方法均以群落的树种数和每个树种的个体数相结合来表达树种的多样性,为进一步分析各树种多样性测度指标出现上述差异的原因,选择样地6,7,8,10和11共5种林分类型的树种株数分布和多样性指标计算过程进行比较(表 4)。
|
|
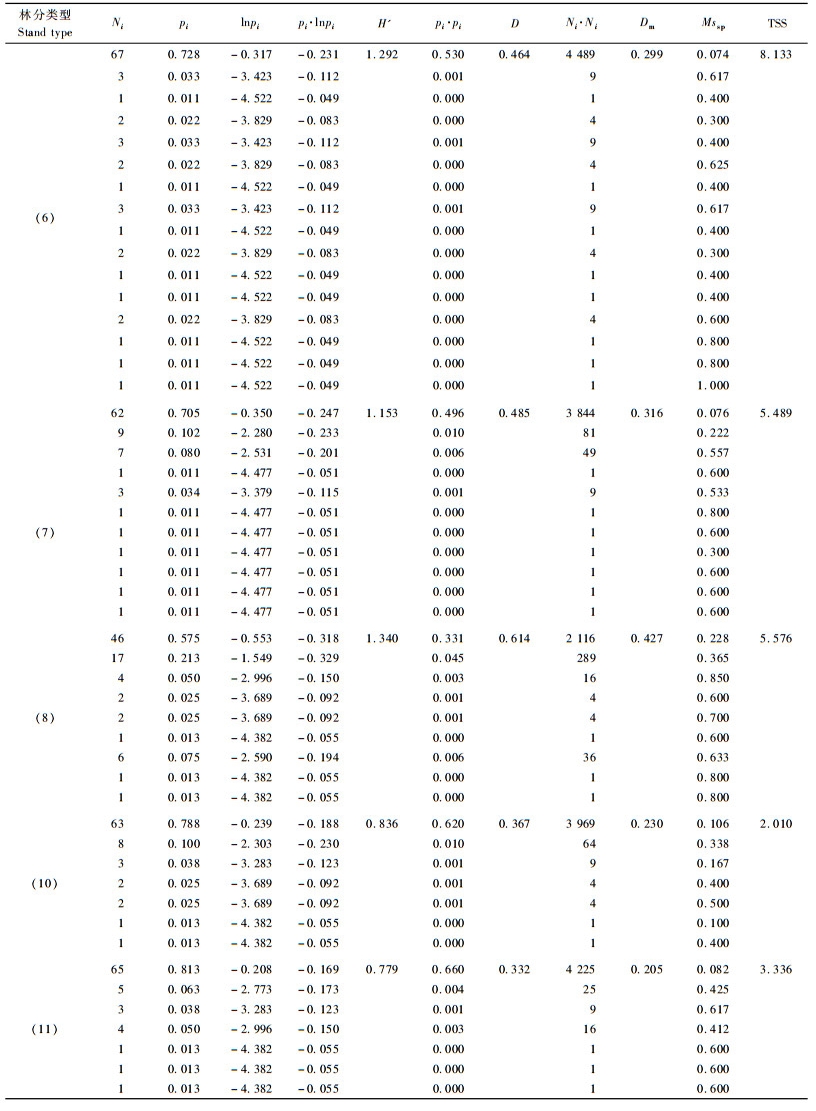
由Shannon-Wiener指数(H′)的计算公式可知,其计算群落多样性时与树种的个体比例密切相关,但其并不是一个严格递增的函数,即当群落中某个种的比例pi小于0.368时,其对群落的H′值的贡献将随pi的增加而增大;当pi大于0.368时,其对群落的H′值的贡献随pi增加而减小(岳天祥,1999;2001)。由表 4可以看出,5个林分类型中的建群种的比例最少为57.5%,随着建群种比例的升高,其对群落多样性的贡献降低。因此,当群落中的树种数一定时,各树种株数分布比例均匀,群落的多样性高,林分类型10的树种多样性高于林分类型11说明Shannon-Wiener指数这一特点,这也是造成林分类型8的多样性高于林分类型6和林分类型7的原因。此外,由表 4的计算过程可以看出,群落中某个种的比例很低时,它对Shannon-Wiener指数值的贡献是很小的,这说明Shannon-Wiener指数对稀少种并不敏感。同样对Simpson指数(D)和MeIntosh指数(Dm)来说,也只考虑了种在群落中株数比例,对稀少种对多样性的贡献不敏感。TSS指数将群落中种的比例和种间的隔离程度相结合来表达树种空间多样性。由表 4的计算过程可以看出,建群种在群落中所占的比例越大,其对多样性指数值的贡献越小,说明群落的树种空间多样性越小;林分类型10和林分类型11物种数均为7个,但林分类型11中稀有种为3个,而林分类型10中为2个,TSS计算结果表明:林分类型11的多样性较林分类型10的高,而与Shannon-Wiener指数计算的结果相反,这说明TSS指数能够明确体现出稀少种对群落多样性的贡献,即群落中树种数一定的情况下,群落中稀少种的种类越多,群落的多样性越大。
4 结论与讨论生物多样性研究是生态系统研究中的一个非常重要的领域。虽然物种多样性的定量研究在不断深入,但目前的物种多样性计算大多还局限于物种丰富度和相对多度的结合,使用不同的多样性模型往往会得出不同的结论。本研究中,模拟数据计算结果表明H′和D不会因为调查数据的多寡而影响到多样性指数的结果,而Dm和α则会受到调查株数和多度的影响,α在相同的调查株数和相同的树种数量时,表现出相同的多样性。实测数据的计算结果也说明上述多样性指数存在的不足。以物种的丰富度和相对多度相结合计算物种多样性,会出现物种数多,多样性反而低的现象,这是由于样地中各树种所占的比例不同而引起的;实测数据同时也表明H′和D对群落中稀有种并不敏感,没有表现出在相同树种数的情况下,稀有种越多,多样性越大的自然规律。模拟数据和实测数据都表明:基于相邻木空间关系的树种空间多样性指数(TSS)具有多样性随物种丰富度增大而增大的一般属性,不受调查株数和样地大小的影响,能够反映稀有种的影响,且该指数能够正确表达出相同物种数量群落的物种空间隔离程度的高低顺序,这是其他指数不具备的特征。在本文中,实测数据使用了固定样地调查和抽样调查2种方法,虽然很好地支撑了以上结论,但也会引起是否由于试验设计和取样方法的原因对以上结论产生疑问,在今后的研究中将进一步收集实测数据,验证该指数的特性。此外,由于该指数是针对树种多样性的测度指数,这在一定程度上适用范围较其他多样性指数窄。即使能够推广应用于群落的草本层或灌木层多样性测试,也可能会因为调查成本上升而影响到其应用,目前尚未有针对草本层和灌木层的应用实例,但对于测度和了解森林经营中树种的多样性变化来说,完全可以运用该指数进行测度。
当前,维持物种多样性已成为森林可持续经营的一个主要内容和重要目标(雷相东等,2000;Hurlbert, 1971),对生物多样性的测度不仅要包括群落中所有物种的多度和丰富度,还应该反映群落之间的生物学上或生态学上的差异,对于森林经营者来说,需要了解森林经营活动前后生物多样性的变化,从而对经营活动做出正确决策和规划,因此,对于生物多样性测度指标的选择就显得十分重要。研究物种水平上的多样性简单从物种的丰富度和各物种的相对多度来结合进行测度显然还不能满足森林经营工作的需要。因为仅从不同林分类型的树种丰富度和各树种相对多度还不能表达森林在结构上的变异,也很难体现经营活动对群落多样性的影响。基于相邻木空间关系的树种空间多样性指数(TSS)作为一种新的树种多样性测度指标,它将群落中树种间的空间隔离关系考虑在内,在一定程度上反映了森林结构的差异,且该指数容易测定,调查成本相对较低,不受样地大小和数量的限制,可以与经营相关的因子联系起来,是一个测度树种多样性的良好指数,也为森林结构多样性指标的构建提供一个很好的参考。
| [] | 惠刚盈, von GadowK, 胡艳波, 等. 2007. 结构化森林经营. 北京, 中国林业出版社: 29-32. |
| [] | 雷相东, 唐守正. 2000. 森林经营对群落α多样性的影响定量研究进展. 生态学杂志, 19(3): 46–51. |
| [] | 马克平. 1993. 试论生物多样性的概念. 生物多样性, 1(1): 20–22. DOI:10.17520/biods.1993005 |
| [] | 马克平. 1994. 生物多样性的测度方法: Ⅰ. α多样性的测度方法(上). 生物多样性, 2(3): 162–168. |
| [] | 马克平, 刘玉明. 1994. 生物多样性的测度方法: Ⅰ. α多样性的测度方法(下). 生物多样性, 2(4): 231–239. |
| [] | 王永繁, 余世孝, 刘尉秋. 2002. 物种多样性指数及其分形分析. 植物生态学报, 26(4): 391–395. |
| [] | 徐海, 惠刚盈, 胡艳波, 等. 2007. 天然红松阔叶林林木分布格局研究的最小样本量. 林业科学研究, 20(2): 160–164. |
| [] | 岳天祥. 2001. 生物多样性研究及其问题. 生态学报, 21(3): 462–467. |
| [] | 岳天祥. 1999. 生物多样性模型研究. 自然资源学报, 14(4): 377–380. DOI:10.11849/zrzyxb.1999.04.017 |
| [] | 赵士洞, 汪业勖. 1997. 生态系统管理的基本问题. 生态学杂志, 16(4): 35–38. |
| [] | Fisher R A, Corbet A S, Williams C B. 1943. The relation between the number of species and the number of individuals in a random sample of an animal population. Animal Ecology, 12(1): 42–58. DOI:10.2307/1411 |
| [] | Ganeshaiah K N, Shaanker U R. 2000. Measuring biological heterogeneity of forest vegetation types: avalanche index as an estimate of biological diversity. Biodiversity and Conservation, 9(7): 953–963. DOI:10.1023/A:1008910918751 |
| [] | Hui G Y, Zhao X H, Zhao Z H, et al. 2011. Evaluation tree species spatial diversity based on neighborhood relationship. Forest Science, 57(4): 292–300. |
| [] | Hurlbert S H. 1971. The non-concept of species diversity: a criteria and alternative parameters. Ecology, 52(4): 577–586. DOI:10.2307/1934145 |
| [] | Margalef R. 1958. Information theory in ecology. General Systematies, 3: 36–71. |
| [] | McIntosh R P. 1967. An index of diversity and the relation of certain concepts to diversity. Ecology, 48(3): 329–404. |
| [] | Magurran A E. 1988. Ecological diversity and its measurement. New Jersey, Princeton University Press: 192. |
| [] | Pielon E C. 1975. Ecological diversity. New York, John Wiley and Sons: 165. |
| [] | Simpson E H. 1949. Measurement of diversity. Nature, 163: 688. DOI:10.1038/163688a0 |
| [] | Shannon C E, Weaver W. 1963. The mathematical theory of communication. Urbana, University of Illinois Press: 117. |
| [] | Whittaker R H. 1972. Evolution and measurement of species diversity. Plant Taxon, 21(3): 213–251. |