 2012, Vol. 48
2012, Vol. 48文章信息
- 徐庆华, 臧润国, 丁易, 许正, 刁永强, 孙霞
- Xu Qinghua, Zang Runguo, Ding Yi, Xu Zheng, Diao Yongqiang, Sun xia
- 天山北坡7种野生果树幼苗的功能性状分析
- Analysis on Seedling Functional Traits of 7 Wild Fruit Tree Species on the Northern Slope of Tianshan Mountains
- 林业科学, 2012, 48(10): 63-69.
- Scientia Silvae Sinicae, 2012, 48(10): 63-69.
-
文章历史
- 收稿日期:2011-11-29
- 修回日期:2012-05-05
-
作者相关文章



2. 新疆伊犁州林业科学研究院 伊宁 835000;
3. 新疆农业大学 乌鲁木齐 830052
2. Yili Academy of Forestry Yining 835000;
3. Xinjiang Agriculture University Urumqi 830052
植物功能性状是近年来生态学研究的一个重要方向(Reich et al., 1999;Wright et al., 2004;张林等,2004;He et al., 2006),植物功能性状通常指影响植物存活、生长、繁殖速率和最终适合度的生物学特征(Ackerly,2003),一般包括植物的生长型、潜在高度、木材密度、比叶面积(specific leaf area, SLA)、固氮能力、叶片氮磷含量和落叶与否等形态、生理和物候等特征(Cornelissen et al., 2003)。功能性状的研究方法能有效利用植物的生理、形态和生活史等特征来反映植物个体、种群、群落和生态系统水平上的生物之间、生物与环境之间的相互作用,同时能够揭示生物对生态系统功能的影响(Diaz et al., 2004;Westoby et al., 2006;Poorter et al., 2008;Hillebrand et al., 2009;Reiss et al., 2009;Zhang et al., 2011)。研究发现,植物可以调整自身的特征属性,从而使其适应较为复杂的环境条件,增加物种在不同环境下的分布范围(Hutchings et al., 1994),与植物生长相关的功能性状表现为趋同适应,而与植物更新和扩散有关的性状则表现为趋异适应,其原因是生物竞争和环境筛同时作用于群落,对共同种的生态策略及生态功能具有不同的效应(Ackerly,2007;Suding et al., 2008)。植物功能性状的研究对环境资源状况评价和植物栽培学研究也起到积极的推动作用,Koerselman等(1996)、Gusewell等(2002)研究发现植物体内的氮磷比是一个具有重要生态意义的功能性状指标,一方面可以反映植物种的结构和功能特征,同时也反映群落水平的营养限制状况,当生境中磷过剩而氮不足时,植物过量吸收磷,植物叶片的氮磷比通常小于14,植物生长表现为氮限制;反之,当植物叶片的氮磷比较大时(一般大于20),植物生长表现为磷限制,对植物增施生长限制养分量可以显著增加植物的生物量。
在天山北坡海拔1 000~1 600 m的中山带阴坡、半阴坡,分布着由新疆野苹果(Malus sieversii)、樱桃李(Prunus cerasifera)、杏(Armeniaca vulgaris)及胡桃(Juglans regia)等组成的野果林,这是在中亚荒漠地带山地罕见的“海洋性”阔叶林类型,是第三纪暖温带阔叶林的孑遗群落,也是北方落叶果树遗传育种十分珍贵的基因库(张新时,1973)。新疆天山北坡特殊的地理、气候环境使新疆野苹果、胡桃、野樱桃等这些在其他地区早已消失的第三纪植物区系起源发展、进化的种群得以保留,这些植物种不仅具有较高的经济价值,而且对于研究中国植物区系进化过程等都具有很高的科研价值。其中新疆野苹果被认为是现代苹果的起源中心之一(张钊,1982;王磊,1989),是第三纪孑遗植物之一(傅立国,1992),已被列为我国具有国际意义的生物多样性优先保护物种和中国渐危二级重点保护植物。但由于近年来掠夺式的资源开发及农业建设等因素,导致新疆野生果树资源破坏严重,野果林面积急剧减少,濒临灭绝。以新疆野苹果为例,1959年新疆野苹果调查统计资料表明总面积曾达9 300 hm2(林培钧等,2000),2006年统计资料显示新疆野苹果总面积仅为5 000 hm2(冯涛等,2006)。因此开展天山北坡野生果树种群的生态科学研究和保护工作刻不容缓。
幼苗是森林生态系统中的一个重要组成部分,它们在物种多样性的维持、群落演替和树种更新等过程中发挥着重要作用(Swamy et al., 2011),幼苗的功能性状反映了幼苗对环境条件的适应策略(路兴慧等,2011)。本文通过对天山北坡主要野生果树幼苗的功能性状特征进行测定,补充和完善植物功能性状基础数据库。了解天山北坡主要野生果树幼苗的功能性状,为进一步深入分析野果林的生态功能奠定基础,为野生果树的资源保护、栽培扩繁提供科学依据。
1 试验区概况试验区位于新疆伊犁新源县东南的野果林改良场,地理位置为43°10′—43°30′ N;82°20′—82°50′ E,海拔1 200~1 600 m,土壤类型为山地黑棕色土,气候为温带大陆性气候,年均温4~8 ℃,1月平均气温为3.0 ℃, 7月平均气温为23~28 ℃,无霜期约130天, 积雪期约150天, 年日照时数2 500 h以上, >10 ℃积温3 000 ℃左右,年降水量550~700 mm。试验样品采集区的野果林面积大约80 hm2,各种野生果树混生,其中新疆野苹果分布面积最大,约占总面积的40%,其次是樱桃李和杏,分别占总面积的10%左右,其他野生果树分布面积相对较少,林内混生有天山茶藨子(Ribes meyeri)、新疆忍冬(Lonicera tatarica)等其他野生植物种。
2 研究方法于2011年7月在试验区野果林内选择新疆野苹果、胡桃、杏、辽宁山楂(Crataegus sanguinea)、樱桃李、酸枣(Zizyphus jujuba var. spinosa)、新疆桃(Amygdalus ferganensis)等7种野生果树的生长健康、高度在40~60 cm的自然生幼苗各20株,并逐一挂牌标记,分别标记幼苗的根、茎、叶取样,测定其地上部分功能性状形态指标和各器官养分含量,地上部分功能性状形态指标测定设置重复20株,并每株选取2片叶子进行扫描处理,根、茎、叶养分测定设置5株重复,所有试验植株地上器官取样都选择在植株向阳面的同一方向枝条上。
测定的植物功能性状形态指标包括:叶片面积、叶片厚度、叶片干物质量、茎皮厚度、木材密度,养分含量指标包括:叶、茎、根的全N、全P、全K含量,根据叶片面积和干物质量计算比叶面积。叶片面积利用扫描仪(型号Canon-scan Lide 100)扫描计算;叶片厚度和茎皮厚度利用游标卡尺测定;叶片干物质量测定是:先将采集叶片阴凉处晾干带回实验室,在80 ℃烘箱内烘干至恒量,天平称量(型号为Sartorius Bs323.s,max.320 g,d=0.001);木材密度测定是利用浸水法测定体积,样品在75~80 ℃烘箱烘干2~3天至恒量,称量法测定质量,计算密度。试验测定方法参考方精云等(2009)植物群落清查技术规范。为测定植物器官养分含量,将带回实验室的植物样品在80 ℃恒温下烘干、粉碎,分别称取样品,经浓硫酸消煮后,全N含量采用凯氏定氮法,全P含量采用钒钼黄比色法,全K含量采用火焰光度计法分别测定记录。
3 结果与分析 3.1 7种野生果树苗木的形态特征比较由表 1可知,不同树种间功能性状的形态指标差异显著(P<0.05),新疆野苹果和杏的叶片厚度较大,均为(0.23±0.01)mm,两者均与胡桃的叶片厚度差异不显著(P>0.05),但均显著大于其他树种的叶片厚度,酸枣和新疆桃的叶片厚度较小,两者差异不显著。但均显著小于其他树种的叶片厚度。虽然酸枣和新疆桃的叶片厚度较小,但两者的比叶面积却较大,除新疆野苹果比叶面积与两者差异不显著外,其他野生果树的比叶面积均显著小于酸枣和新疆桃,胡桃的比叶面积最低,显著小于其他野生果树(P<0.05)。
|
|

茎皮厚度从大到小的顺序依次为胡桃、辽宁山楂、杏、新疆桃、樱桃李、新疆野苹果和酸枣,其中胡桃、辽宁山楂和杏的茎皮厚度差异不显著(P>0.05),酸枣的茎皮厚度均显著小于其他树种。酸枣木材密度最高,为(0.79±0.03)mg·mm-3,新疆野苹果的木材密度最小,为(0.53±0.02)mg·mm-3。除樱桃李与酸枣的木材密度差异不显著外,其他树种木材密度均显著小于酸枣(P<0.05)。
3.2 7种野生果树苗木不同器官养分含量比较 3.2.1 叶片养分含量比较由表 2可知,各种野生果树的叶N含量差异显著(P<0.05),各树种叶N含量均在20 mg·g-1以上,酸枣叶片N含量最高,为(37.6±2.0)mg·g-1,辽宁山楂最低,为(20.7±0.7)mg·g-1。酸枣、新疆桃和杏的叶N含量相互之间差异不显著,樱桃李、胡桃和辽宁山楂叶N含量相互之间差异不显著(P>0.05),各树种叶N含量大小顺序依次是酸枣>新疆桃>杏>新疆野苹果>樱桃李>胡桃>辽宁山楂。各种野生果树的叶P含量差异显著(P<0.05),樱桃李叶P含量最高,为(4.1±0.2)mg·g-1,胡桃、杏和酸枣的叶P含量较低,为(1.5±0.2)mg·g-1左右,其他树种居中。各种野生果树的叶K含量差异不显著(P>0.05),新疆野苹果、杏和樱桃李的叶K含量较高,均为(9.4±0.2)mg·g-1左右,胡桃叶K含量最低,为(8.8±0.2)mg·g-1。7个树种的叶N含量平均为28.4 mg·g-1,叶P含量平均为2.2 mg·g-1,叶K含量平均为9.2 mg·g-1左右。
|
|
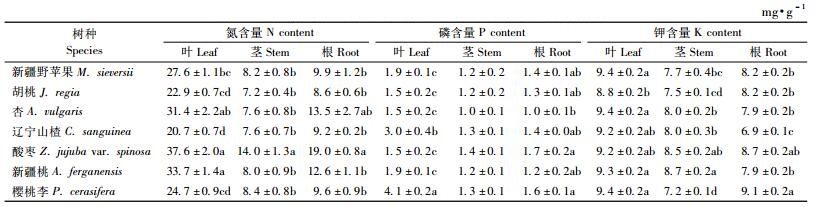
由表 2可知,酸枣茎N含量最高,平均为(14.0±1.3)mg·g-1,显著高于其他树种茎部N含量(P<0.05),胡桃茎N含量最低,为(7.2±0.4)mg· g-1,除酸枣外,其他树种茎部N含量差异不明显(P>0.05)。各树种的茎部P含量差异不显著(P>0.05),平均都在1.4~1.0 mg·g-1。各树种的茎部K含量差异显著(P<0.05), 新疆桃茎部K含量最高,为(8.7±0.2)mg·g-1,樱桃李的茎K含量最低,为(7.2±0.1)mg·g-1。
3.2.3 根养分含量比较由表 2可知,各种野生果树的根N含量差异显著(P<0.05),杏、酸枣和新疆桃的根N含量在12~19 mg·g-1,其他树种根N含量均在10 mg·g-1以下,酸枣根N含量最高,为(19.0±0.8)mg·g-1,胡桃的根N含量最低,为(8.6±0.6)mg·g-1。各种野生果树的根部P含量差异显著(P<0.05),均在1.0~1.7 mg·g-1,与根N相比含量较低。除杏外,其他树种的根P含量两两之间均差异不显著(P>0.05)。各种野生果树的根K含量差异显著(P<0.05),各种野生果树的根K含量在6.9~9.1 mg·g-1,野樱桃的根K含量最高,为(9.1±0.2)mg·g-1,除与酸枣差异不显著外(P>0.05),均显著大于其他树种的根K含量(P<0.05),辽宁山楂最低,为(6.9±0.1)mg·g-1,均显著小于其他树种的根K含量(P<0.05)。
3.3 7种野生果树苗木根、茎、叶器官N/P比值差异图 1表明:不同树种叶片N/P比值差异显著(P<0.05),酸枣与杏的叶片N/P比值较大,分别约为24.9和20.9,均大于20;辽宁山楂和樱桃李的叶片N/P比值较低,分别为6.9和,6.0,均小于10,显著低于其他树种;新疆野苹果、胡桃和新疆桃的叶片N/P比值之间差异均不显著,均在14~20。茎和根器官的N/P比值均小于15,不同树种茎的N/P比值差异不显著。杏根的N/P比值与酸枣和新疆桃差异不显著,但与其他树种根差异显著(P<0.05)。除杏外,其他树种的根N/P比值相互之间均差异不显著(P>0.05)。

|
图 1 不同树种各器官的N/P值比较 Fig.1 The organic N/P ratios in different trees species |
通过对7个树种的比叶面积(SLA)与其叶片N,P含量进行Pearson相关性检验表明:比叶面积(SLA)与叶片N含量呈显著正相关(R=0.874*;P=0.01),与叶片P含量相关性不显著(R=-0.334;P=0.464),比叶面积与叶片N含量相关关系见图 2。

|
图 2 比叶面积(SLA)与叶片氮含量相关关系 Fig.2 The linear relationship between SLA and leaf N |
植物比叶面积(SLA)的差异,往往反映不同物种的生长策略分化,从而能在同一环境内稳定共存。树种的SLA越高,吸收光照资源的能力越强,越利于生长,但对叶片的防御保护不够,容易被食叶动物啃食(Rossatto et al., 2009)。在植物种长期进化过程中,不同物种的生长型、生长速率、资源获取等生态策略和生活史方面都表现出差异性,从而减少物种间竞争,促进物种共存;另外,树种的木材密度与植物水分传导呈负相关,与水分利用效率和耐干旱能力呈正相关,茎皮厚度与抗病虫害和抗旱能力相关(Cornelissen et al., 2003;Marketstejin et al., 2007;Cornwell et al., 2009)。本试验所测定的7种野生果树苗木比叶面积(SLA)差异反映了不同树种能够在同一环境中长期共存。分析表明SLA与其叶片N含量呈显著正相关,Wright等(2004)对全球175个地区2 548植物种的研究认为,叶片N含量越高,植物SLA越大,光合作用越强,叶片生产力越高,其他相关研究也验证了类似的结论(Poorter,1999;Rossatto et al., 2009)。从木材密度和茎皮厚度来看,酸枣、樱桃李木材密度较大,可能两者的水分利用效率较其他野生果树高,耐旱性能力较强些,这也可能与其生长速度较慢有关;Osunkoya等(2007)认为物种的木材密度越低,其生长速率越高,但导致其树体结构不稳定和耐荫性差;胡桃、辽宁山楂和杏茎皮厚度较大,表明其抵抗病虫害等不良环境因素能力较强,新疆野苹果和酸枣茎皮厚度较小,表明其抵抗病虫害等不良环境因素能力较弱。近来伊犁新源地区新疆野苹果林出现大面积枯死现象,枝干被蛀干害虫侵入,可能与其茎皮厚度和木材密度小有关(成克武等,2008)。
植物叶片N,P含量与N/P比值特征研究(Koerselman et al., 1996;Thompson et al., 1997;Braakhekke et al., 1999;Elser et al., 2000;Gusewell,2004)表明:叶片的N,P含量和N/P比值反映植物对当地土壤条件的适应,叶片N,P含量与光合能力和生长呈正相关,与叶片寿命呈负相关。Gusewell等(2002)研究陆地生态系统植物叶片N/P的阈值认为,当N/P < 10时,增加N肥可以显著增加植物的生物量,当N/P>20,增加P肥可以显著增加植物的生物量,N/P比值在两者之间时,施肥对生物量的效果与N/P比值关系不明显。本试验测定的杏和酸枣叶N/P值均大于20,辽宁山楂和樱桃李叶N/P比值均小于10,其他树种叶N/P比值均在10~20。因此,在进行天山北坡的野生果树栽培时,对土壤增施N肥可能较显著地增加辽宁山楂和樱桃李的生物量,对土壤增施P肥可能较显著地增加酸枣和杏的生物量。
Elser等(2000)对全球陆地植物研究发现,叶片N含量平均为20.6 mg·g-1,P含量平均为1.99 mg·g-1;Han等(2005)对中国陆地植物的研究发现,叶片N含量平均为20.2 mg·g-1,P含量平均为1.46 mg·g-1,叶片N含量与全球水平差不多,而P含量低于全球平均水平,认为中国陆地土壤与全球相比,P含量偏低。由表 2可知,天山北坡7种主要野果树种叶N含量平均为28.4 mg·g-1,P含量平均为2.2 mg·g-1,也均高于全国和全球陆地植物叶片N,P含量的平均水平。由此可见,在天山北坡7种试验野生果树的自然分布区内,土壤N,P养分含量均高于国内外陆地土壤N,P养分含量平均水平。也有学者认为植物叶片较高的N,P含量是植物为抵消低温对代谢反应抑制的一种适应机制(Reich et al., 2004),由此来看,7种野生果树叶片较高的N,P含量也可能是对其生长在低温环境的一种功能性适应。
养分利用效率通常是指吸收单位数量养分所能生产的干物质量,根据目前应用较多的Chapin指数法(Chapin,1980;Birk et al., 1986;Safou-Matondo et al., 2005;Ducic et al., 2009),以植物生物量与植物养分量的比值作为养分利用效率,即利用植物器官或组织养分含量的倒数作为养分利用效率指标,指标值越大养分利用效率越高,指标值越小养分利用效率越低。根据植株苗木根、茎、叶器官的N,P,K总含量,酸枣根,茎,叶的N,P,K总含量分别为29.4,23.9,48.3 mg·g-1,均为试验测定树种中最高,新疆桃仅次于酸枣,根、茎、叶的N,P,K总含量分别为21.7,17.9,44.9 mg·g-1,辽宁山楂和胡桃根、茎、叶的N,P,K养分总含量较少,其他树种居中。这些数据在一定程度上说明新疆桃和酸枣喜欢生长在较肥沃的土壤上,耐贫瘠能力较差,辽宁山楂和胡桃能够在养分贫乏的土壤上生长,耐贫瘠能力较强,其他树种居中。据此,在野生树种栽培时,新疆桃和酸枣应选择较为肥沃的土地,养分较为贫乏的土壤尽可能栽培辽宁山楂和胡桃。虽然树种生长对养分需求的差异需要进一步试验分析,但从一个侧面反映了野生果树生长对土壤养分要求和适应的功能性差异。根据养分利用效率,可以把所研究的7个树种分为3类:新疆桃和酸枣属于养分利用率较低型,辽宁山楂和胡桃的养分利用率较高型,其他3个树种属于养分利用效率中等型。
| [] | 成克武, 周晓芳, 臧润国, 等. 2008. 新疆野苹果资源保护对策探讨. 干旱区研究, 25(6): 760–765. |
| [] | 方精云, 王襄平, 沈泽昊, 等. 2009. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 17(6): 533–548. |
| [] | 冯涛, 张红, 陈学森, 等. 2006. 新疆野苹果果实形态与矿质元素含量多样性以及特异性状单株. 植物遗传资源学报, 7(3): 270–276. |
| [] | 傅立国. 1992. 中国植物红皮书. 北京, 科学出版社. |
| [] | 林培钧, 崔乃然. 2000. 天山野果林资源——伊犁野果林综合研究. 北京, 中国林业出版社. |
| [] | 路兴慧, 丁易, 臧润国, 等. 2011. 海南岛热带低地雨林老龄林木本植物幼苗的功能性状分析. 植物生态学报, 35(12): 1300–1309. |
| [] | 王磊. 1989. 新疆野苹果和新疆野杏. 新疆农业科学, 31(6): 33–34. |
| [] | 张林, 罗天祥. 2004. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 28(6): 844–852. DOI:10.17521/cjpe.2004.0110 |
| [] | 张新时. 1973. 伊犁野果林的生态地理特征和群落学问题. 植物学报, 15(2): 239–253. |
| [] | 张钊. 1982. 新疆野苹果. 乌鲁木齐, 新疆人民出版社. |
| [] | Ackerly D D. 2003. Community assembly, niche conservation, and adaptive evolution in changing environments. International Journal of Plant Science, 164(3): 165–184. |
| [] | Ackerly D D. 2007. A trait-based approach to community assembly:partitioning of species trait values into within-and among-community components. Ecology Letters, 10(2): 135–145. DOI:10.1111/ele.2007.10.issue-2 |
| [] | Birk E M, Vitousek P M. 1986. Nitrogen availability and nitrogen use efficiency in loblolly pine stands. Ecology, 67(1): 69–79. DOI:10.2307/1938504 |
| [] | Braaldaekke W G, Hooftman D A P. 1999. The resource balance hypothesis of plant species diversity in grassland. Journal of Vegetation Science, 10(2): 187–200. DOI:10.2307/3237140 |
| [] | Chapin F S. 1980. The mineral nutrition of wild plants. Annual Review of Ecology and Systematics, 48(11): 233–260. |
| [] | Cornelissen J H C, Lavorel S, Garnier E, et al. 2003. A Handbook of protocols for standardized and easy measurement of plant functional traits world-wide. Australian Journal of Botany, 51(4): 335–380. DOI:10.1071/BT02124 |
| [] | Cornwell W K, Ackerly D D. 2009. Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California. Ecological Monographs, 79(1): 109–126. DOI:10.1890/07-1134.1 |
| [] | Diaz S, Hodgson J G, Thompson K, et al. 2004. The plant traits that drive ecosystems:evidence from three continents. Jounal of Vegetation Science, 15(3): 295–304. DOI:10.1111/j.1654-1103.2004.tb02266.x |
| [] | Ducic T, Berthold D, Langenfeld-Heyser R, et al. 2009. Mycorrhizal communities in relation to biomass production and nutrient use efficiency in two varieties of Douglas fir (Pseudotsuga menziesii var. menziesii and var. glauca) in different forest soils. Soil Biology and Biochemistry, 41(4): 742–753. DOI:10.1016/j.soilbio.2009.01.013 |
| [] | Elser J J, Fagan W F, Denno R F, et al. 2000. Nutritional constraints in terrestrial and freshwater food webs. Nature, 408: 578–580. DOI:10.1038/35046058 |
| [] | Gusewell S, Koerselman M. 2002. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology Evolution and Systematics, 5(1): 37–61. DOI:10.1078/1433-8319-0000022 |
| [] | Gusewell S. 2004. N/P ratios in terrestrial plants:variation and functional significance. New Phytologist, 164(2): 243–266. DOI:10.1111/j.1469-8137.2004.01192.x |
| [] | Han W X, Fang J Y, Guo D L, et al. 2005. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 168(1): 377–385. |
| [] | He J S, Wang Z H, Wang X P. 2006. A test of the generality of leaf trait relationship on the Tibetan Plateau. New Phytologist, 170(4): 835–848. DOI:10.1111/nph.2006.170.issue-4 |
| [] | Hillebrand H, Matthiessen B. 2009. Biodiversity in a complex world:consolidation and progress in functional biodiversity research. Ecology Letters, 12(12): 1405–1419. DOI:10.1111/j.1461-0248.2009.01388.x |
| [] | Hutchings M J, Kroon D H. 1994. Foraging in Plants:the role of morphological plasticity in resource acquisition. Advance in Ecological Research, 25(1): 159–238. |
| [] | Koerselman W, Meuleman A M. 1996. The vegetation N:P ratio:a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 33(6): 1441–1450. DOI:10.2307/2404783 |
| [] | Marketstejin L, Poorter L, Bongers F. 2007. Light-dependent leaf trait variation in 43 tropical dry forest tree species. American Journal Botany, 94(4): 515–525. DOI:10.3732/ajb.94.4.515 |
| [] | Osunkoya O O, Sheng T K, Mahmud N A, et al. 2007. Variation in wood density, wood water content, stem growth and mortality among twenty-seven tree species in a tropical rainforest on Borneo Island. Austral Ecology, 32(2): 191–201. DOI:10.1111/aec.2007.32.issue-2 |
| [] | Poorter L. 1999. Growth responses of 15 rain-forest tree species to a light gradient:the relative importance of morphological and physiological traits. Functional Ecology of Plants, 13(3): 396–410. DOI:10.1046/j.1365-2435.1999.00332.x |
| [] | Poorter L, Wright S J, Paz H, et al. 2008. Are function traits good predictors of demographic rates?Evidence from five neotropcial forests. Ecology, 89(7): 1908–1920. DOI:10.1890/07-0207.1 |
| [] | Reich P B, Ellsworth D S, Waiters M B, et al. 1999. Generality of leaf trait relationships:a test across six biomes. Ecology, 80(6): 1955–1969. DOI:10.1890/0012-9658(1999)080[1955:GOLTRA]2.0.CO;2 |
| [] | Reich P B, Oleksyn J. 2004. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 101(30): 11001–11006. DOI:10.1073/pnas.0403588101 |
| [] | Reiss J, Bridle J R, Montoya J M, et al. 2009. Emerging horizons in biodiversity and ecosystem functioning research. Trends in Ecology and Evolution, 24(9): 505–514. DOI:10.1016/j.tree.2009.03.018 |
| [] | Rossatto D R, Hoffman W A, Franco A C. 2009. Differences in growth patternsco-occurring forest and savanna trees affect the forest-savanna boundary. Functional Ecolgy, 23(4): 689–698. DOI:10.1111/j.1365-2435.2009.01568.x |
| [] | Safou-Matondo R, Deleporte P, Laclau J P, et al. 2005. Hybrid and clonal variability of nutrient content and nutrient use efficiency in Eucalyptus stands in Congo. Forest Ecology and Management, 210(3): 193–204. |
| [] | Suding K N, Lavorel S, Chapin Ⅲ F S, et al. 2008. Scaling environmental change through the community level:a trait based response and effect framework for plants. Global Change Biology, 14(5): 1125–1140. DOI:10.1111/j.1365-2486.2008.01557.x |
| [] | Swamy V, Terborgh J, Dexter KG, et al. 2011. Are all seeds equal?Spatially explicit comparisons of seed fall and spring recruitment in a tropical forest. Ecology Letters, 14(2): 195–201. DOI:10.1111/j.1461-0248.2010.01571.x |
| [] | Thompson K, Parkinson J A, Band S R, et al. 1997. A comparative study of leaf nutrient concentrations in a regional herbaceous flora. New Phytologist, 136(4): 679–689. DOI:10.1046/j.1469-8137.1997.00787.x |
| [] | Westoby M, Wright I J. 2006. Land-plant ecology on the basis of functional traits. Trends in Ecology and Evolution, 21(5): 261–268. DOI:10.1016/j.tree.2006.02.004 |
| [] | Wright I J, Reich P B, Westoby M, et al. 2004. The worldwide leaf economics spectrum. Nature, 428: 821–827. DOI:10.1038/nature02403 |
| [] | Zhang S B, Slik J F, Zhang J L, et al. 2011. Spatial patterns of wood traits in China are controlled by phylogeny and the environment. Global Ecology and Biogeography, 20(2): 241–250. DOI:10.1111/j.1466-8238.2010.00582.x |