 2012, Vol. 48
2012, Vol. 48文章信息
- 齐磊, 胡德夫, 丁长青, 隋金玲, 张东, 杨亮, 吴记贵, 蒋万杰
- Qi Lei, Hu Defu, Ding Changqing, Sui Jinling, Zhang Dong, Yang Liang, Wu Jigui, Jiang Wanjie
- 北京松山国家级自然保护区鼠类群落多样性与结构变动分
- Rats Community Structure and Diversity in the Songshan National Nature Reserve, Beijing
- 林业科学, 2012, 48(9): 181-185.
- Scientia Silvae Sinicae, 2012, 48(9): 181-185.
-
文章历史
- 收稿日期:2011-06-17
- 修回日期:2012-01-09
-
作者相关文章



2. 北京松山国家级自然保护区管理处 北京 102115
2. Songshan National Nature Reserve Management Department Beijing 102115
啮齿动物是哺乳动物中种类最多、数量最多、分布最广的一个类群。在自然界中啮齿动物是一些食肉动物的主要食物来源,是陆地上许多类型生态系统中食物链的重要环节。近年来,国内外对啮齿类群落的研究已有很多报道(张知彬等,2001; 武晓东等,1999; 2000;杨扬等,2008; 周旭东等,2004; Aragon et al., 2009; Courtalon et al., 2010; Tibault etal.,2010)。组成群落的各个物种随气候、小生境和食物的变化,它们种群数量和生活史性状等各因子便会变化。由此群落的物种组成、空间结构、多样性和种间关系等,也随时间而变化(曾宗永,1994)。1987—1989年,陈卫等(1990)对松山保护区鼠类进行过初步研究,时隔20年,北京松山国家级自然保护区启动了综合考察项目。本文针对该项目调查结果中的鼠类群落多样性与结构变动进行了分析与探讨,分析鼠类结构特征和参数,为松山自然保护区生态系统的相关研究提供数据,对选择正确的鼠害防治措施具有重要意义。
1 材料与方法 1.1 研究区域松山国家级自然保护区位于北京市延庆县西北部,地理坐标为115°43'44″—115°50'22″E,40°29'9″—40°33'35″N,总面积为4 671 hm2。保护区属暖温带大陆性季风气候区,与延庆盆地相比,气温偏低、湿度偏高,是典型的山区小气候特征,年平均气温8.5 ℃,最高39 ℃,最低-27.3 ℃,年平均日照2 836.3 h,≥10 ℃的积温3 398.1 ℃,无霜期约150 d,年降水量493 mm,年蒸发量1 772 mm。
松山自然保护区作为华北地区唯一的最大的油松(Pinus tabulaeformis)天然聚集地,区内分布的天然油松林具有很高的保护价值; 此外,保护区内还分布有蒙椴(Tilia mongolica)、核桃楸(Juglans mandshurica)、榆树(Ulmus pumila)、白桦(Betula platyphylla)、黑桦(B. davurica)和大叶白蜡(Fraxinus rhynchophylla)为主的落叶阔叶次生林。保护区内拥有兽类15科29种,爬行类5科15种,两栖类2科2种,鱼类2科12种,特别是鸟类资源丰富,26科119种及6亚种,占北京地区鸟类总数的36.4%(北京市林业局,2011)。由此可见,保护区内的野生动植物资源十分丰富。
1.2 研究方法2009年和2010年的6月和10月采用铗捕法分别对北京松山国家级自然保护区的鼠类齿动物进行了调查,调查区域根据森林植被类型被划分为6个生境类型:(Ⅰ)居民生活区,(Ⅱ)低山灌草从区,(Ⅲ)天然针叶林,(Ⅳ)落叶阔叶林,(Ⅴ)针阔混交林,(Ⅵ)高山灌丛林。铗距5 m,铗线距20 m,共设置样地12个,每个样地不少于100铗,4次共下2 006铗,用生花生为饵,每日下午置铗,24 h收铗。对捕获的鼠标本进行解剖和常规生物学测量,用鼠类种数(S)、捕获率(C)、构成比(G)、多样性指数(H')、均匀性指数(J')、优势度(D)、相似性指数(I)7个指标对不同生境的鼠类群落结构进行统计分析。
群落多样性指数(diversity index H'),采用Shannon-Wiener指数(Margalef,1958)计算:

|
群落均匀性指数(evenness index J'),采用Pielou(1966)均匀度指数计算:
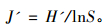
|
群落优势度指数(dominance index) D,采用Simpson(1949)生态优势度指数计算:
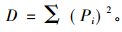
|
群落相似性指数(similarity index) I,采用Whittaker(1960)的相似性指数计算:
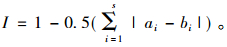
|
式中,S为组成群落的鼠种数,lnS为最大多样性指数,Pi为第i鼠种个体占群落的比例,ai为物种i的个体数在a群落中的比例(%),bi为物种i的个体数在b群落中的比例(%)。
2 结果与分析 2.1 鼠种组成4次调查共捕获鼠类95只,经分类鉴别,分属2科4属8种,分别是鼠科(Muridae)的中华姬鼠(Apodemus draco)(Ad)10只(10.53%),朝鲜姬鼠(A. peninsulae)(Ap)11只(11.58%),小眼姬鼠(A. uralensis)(Au)7只(7.37%),黑线姬鼠(A. agrarius)(Aa)7只(7.37%),社鼠(Niviventer cofueiamis)(Nc)32只(32.68%),褐家鼠(Rattus norvegicus)(Rn)15只(15.79%),小家鼠(Mus musculus)(Mm),12只(12.63%); 仓鼠科(Cricetidae)的大仓鼠(Cricetulustriton)(Ct)1只(1.05%)
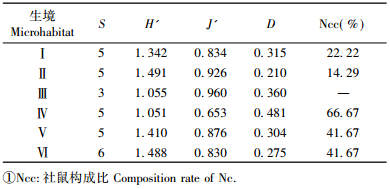
2.2 不同生境各鼠种群构成比较不同生境内鼠类群落构成(种类数和构成比)存在一定差异(表 1) :
|
|
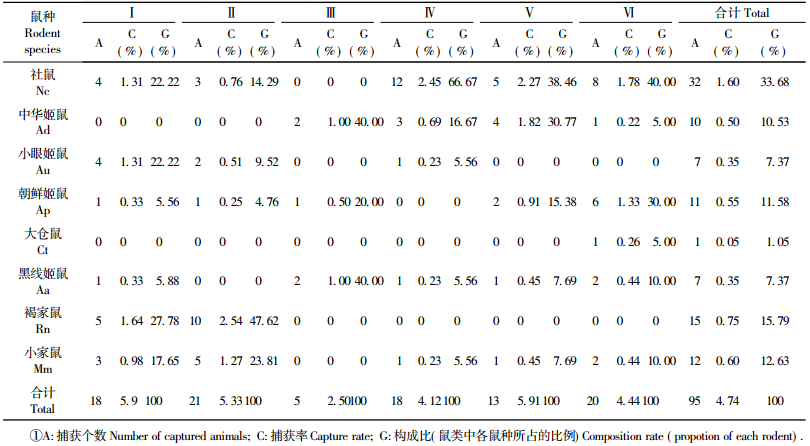
(Ⅰ)居民生活区:海拔800~900 m,人为活动频繁的区域,包括有田地、居住区、餐饮区和办公区及周边乔灌林。主要鼠类是小家鼠和褐家鼠,杂有其他鼠类。
(Ⅱ)低山灌丛区:海拔800 m以下,主要植物有荆条(Vitex negundo var. heterophylla),小叶鼠李(Rhamnus parvifolia)等,有少量人工油松林。鼠类有社鼠、朝鲜姬鼠、小眼姬鼠、褐家鼠、小家鼠。
(Ⅲ)天然次生针叶林:海拔在1 100~1 300 m,树木分布范围狭长,见于塘子沟的西坡和山脊,树种为天然油松。鼠类有中华姬鼠、朝鲜姬鼠、黑线姬鼠。
(Ⅳ)中高山落叶阔叶林: 900~1 300 m,分布在阴坡、半阴坡,密度大。主要乔木为白桦、黑桦、山杨(Populus davidiana)、山杏(Armeniaca sibirica)、蒙椴及核桃楸等,杂有六道木(Abelia biflora)、红丁香(Syringa villosa)等灌木。鼠类有社鼠、中华姬鼠、小眼姬鼠、黑线姬鼠、小家鼠。
(Ⅴ)针阔混交林: 900~1 300 m,多为山势阴坡,较湿润。由兴安落叶松(Larix gmelinii)、油松、桦、槲树(Quercus dentata)及山杨等组成。鼠类有社鼠、中华姬鼠、朝鲜姬鼠、黑线姬鼠、小家鼠。
(Ⅵ)中高山灌丛林:海拔900~1 300 m,多为山势阳面,较干旱。主要植物为红丁香、六道木、胡枝子(Lespedeza bicolor)和平榛(Corylus heterophylla),杂有山杏、核桃楸、秀丽槭(Acer elegantulum)等。鼠类有社鼠、中华姬鼠、朝鲜姬鼠、大仓鼠、黑线姬鼠、小家鼠。其中鼠种类数量Ⅵ最多,有6种; 其次是Ⅰ,Ⅱ,Ⅳ,Ⅴ有5种; Ⅲ最少,为3种。不同生境鼠类捕获率,从高到低依次为Ⅱ,Ⅰ,Ⅴ,Ⅵ,Ⅳ,Ⅲ。由此看出Ⅲ类生境显然不适合该地区大多数鼠类生存和活动。而Ⅰ,Ⅱ类生境,从捕获率上看,是鼠类密度最大的区域。从各鼠种不同生境构成比来看,现松山地区优势种为社鼠、中华姬鼠和朝鲜姬鼠。构成比至少在3个生境中达到15%以上即为优势种,一般用捕获比例大的物种作为优势种(武晓东等,1994; 鲍毅新等,1995)。结果与20年前陈卫等(1990)调查的结果比较,该保护区的黑线姬鼠在鼠类动物中的构成比明显下降,由原来的17.53%减少到现在的7.53%,已不是该地区的优势种,朝鲜姬鼠数量增加,成为优势种。并且本调查新纪录到了小眼姬鼠,收集到的标本符合该鼠种特征,使得松山自然保护区内兽类纪录由29种(陈卫等,1991)增加到30种。
2.3 不同生境群落的多样性、均匀性、优势度比较群落的多样性指数能反映群落物种的丰富性,体现不同群落的异质性。6类生境多样性指数见表 2,由表 2可以看出: Ⅵ类生境群落含鼠种类最多,多样性指数也最高,优势度很低。Ⅰ,Ⅱ和Ⅴ类生境群落的多样性指数、均匀性指数均较高,优势度较低。Ⅳ类生境群落均匀性指数最低,优势度最高。由于在Ⅵ类生境中,自然条件复杂,植被盖度高,食物成分较复杂,为其中鼠类的生存创造了条件,同时受人类活动影响小,所以此类生境内鼠类群落较复杂。Ⅲ类生境中,植被简单,在保护区内形成孤立区域分布且面积较小,因此多样性指数较低。Ⅱ类生境的群落优势度最低,但均匀性和多样性都较高,表明该群落内各鼠之间相互作用,对资源、空间等的利用处于均衡,分布较均匀。分析结果与张洁(1984)对北京地区鼠类群落结构的研究基本一致,因此,在鼠类多样性大的群落中,植物多样性也大; 鼠类多样性小的群落中,植物多样性也小(周庆强等,1982)。
|
|
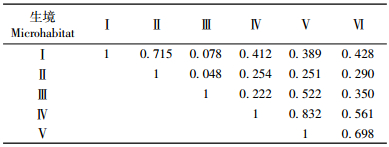
不同生境中鼠类群落相似性分析结果见表 3。根据表 3数据,对6个不同生境的群落相似性指数进行系统聚类分析(图 1)。图 1显示: Ⅲ类生境中鼠类群落与Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅵ类生境中鼠类群落间的相似性均很低,与其植被类型有很大的相关,说明Ⅲ类生境中在保护区内具有特殊的鼠类群落,与其他生境中鼠类群落相比,具有较高的独立性。Ⅳ与Ⅴ类生境中鼠类群落之间的相似性最高(0.832),Ⅰ与Ⅱ次之(0.715),说明Ⅳ与Ⅴ,Ⅰ与Ⅱ的鼠类群落组成接近,优势鼠种相同,结论与表 1数据相吻合。由表 3和图 1可以看出:松山自然保护区内的鼠类群落可以明显的分为3个类群,即Ⅰ-Ⅱ、Ⅲ和Ⅳ-Ⅴ-Ⅵ,体现了依植被类型与海拔变化的鼠类群落间的差异。6个不同生境的鼠类群落相似性指数最后在0.048处聚在一起。结果与高武等(1990)的调查结果相比,保护区内的啮齿类动物经过20年的演替,群落结构已经发生很大变化。
|
|

|
图 1 不同生境啮齿类群落的相似性指数聚类分析 Fig.1 Dendrogram of similarity index of rodent communities in different habitats |
根据调查区域的森林植被类型把松山自然保护区的啮齿类群落生境划分为6个,与传统的依植被带划分相同(周庆强等,1982; 王淯等,1997; 2003;张美文等,2003)。松山自然保护区内植物调查报告显示:较20年前该区域植被类型并未发生变化,所以植被变化对该区域内啮齿动物结构变动的影响甚微(北京市林业局,2011)。调查区域设在坡度大、植被茂密、地势险的山区,所以采取铗线距20 m、小样方多样地进行调查。结果表明:不同生境内的鼠类动物,不论是种群数量还是组成结构都各不相同。说明不同生境内的小气候、植被类型、人为干扰等因素对鼠类动物栖息地选择造成的影响各不相同,食物和隐蔽条件会影响鼠类群落多样性指标。调查结果显示: Ⅱ内的鼠类动物密度最大,Ⅰ次之。Ⅰ类生境是人类活动影响最频繁的区域,地势平缓,在此生境内小家鼠和褐家鼠占群落的50%以上; Ⅱ类生境与Ⅰ类生境接壤,植被茂盛,可使适应的鼠种数量大大的增加,如小眼姬鼠。因此,Ⅰ,Ⅱ区域已接近中度成鼠害(捕获率>6%为中度鼠害)。可以认为此类生境因受人类活动影响,区域内食物(包括饭店贮藏食品、厨余垃圾、游客丢弃食品等)充足,灌木丛和建筑物提供合适的隐蔽场所,造成区域性鼠害发生。关于人类活动对鼠类群落多样性的影响,有研究者认为,人类活动越剧烈,干扰程度越高,多样性与丰富度越低(吴德林等,1993; 张美文等,2003; )。反观松山自然保护区内的鼠类群落多样性和丰富度,显然不符合这一规律。陈志平等(1996)等在对啮齿动物群落多样性的多尺度分析中也提出:啮齿动物群落多样性对尺度变化与不同干扰类型有关。人类对森林的干扰可降低鼠种丰富度和鼠类群落的多样性,这一结论并不是普遍适用,笔者认为:随着空间尺度和干扰类型的不同,鼠类动物抗干扰能力也不相同,群落多样性的变动幅度也会有差异性。
3.2 新纪录小眼姬鼠本次调查新录到小眼姬鼠,并发现现今鼠类动物群落结构与20年前有明显变化。黑线姬鼠构成比明显下降,而其所下降的区域正是小眼姬鼠出现的区域,朝鲜姬鼠结构比例上升。由此可以推论:由于新记录的小眼姬鼠与黑线姬鼠存在种间竞争,经过演变,Ⅰ,Ⅱ类生境内小眼姬鼠占优势,黑线姬鼠下降恰由小眼姬鼠和朝鲜姬鼠所补充。通过对北京林业大学生物学院2006—2010年本科实习用同样方法置铗的记录数据查阅分析,印证了小眼姬鼠的出现对保护区内啮齿动物群落结构有影响,但在种间结构数量上的相应关系需进一步研究分析。
3.3 群落多样性统计分析松山自然保护区内的鼠类动物群落的多样性同物种数呈不显著正相关(r=0.709,P>0.05),其他学者对不同的生态环境研究也得到相同的结果(武晓东等,1994; 陈志平等,1996; 袁帅等,2011)。其次,鼠类动物群落的多样性与均匀性呈不显著正相关(r=0.359,P>0.05),均匀性与群落内优势种所占百分比呈负相关(r=-0.781,P>0.05),Grant等(1979)、周庆强等(1982)、鲍毅新等(1995)、王淯等(1997)、Mangan等(2000)等也得到同样的结果。同时笔者也发现:保护区内的鼠类动物群落的多样性与优势度呈显著负相关(r=-0.883,P<0.05),这与周旭东等(2004)、王淯等(2003)的结果一致,而多样性与其优势种百分比有负相关性和(r=-0.703,P>0.05),王淯等(1997)的结果存在差异。松山自然保护区内的啮齿动物不同生境的群落多样性与群落内优势种比例是否有极显著关系,需要进一步的研究。
综上所述,群落的多样性和均匀性指数随着群落内物种数的增加而增加,但是随着群落内优势种所占比例增加,群落的多样性和均匀性指数都降低。北京松山自然保护区内的啮齿动物具有地带性鼠类群落,会因空间、营养等生态位的不同而引起数量和结构的变化,这是与其他地区研究存在差异的原因。
| [] | 鲍毅新, 丁平, 诸葛阳, 等. 1995. 舟山岛东部地区小型兽类的群落组成与动态分析. 兽类学报, 15(3): 222–228. |
| [] | 北京市林业局. 2011. 松山自然保护区考察专集. 北京, 中国林业出版社. |
| [] | 陈卫, 高武. 1990. 北京松山自然保护区鼠类群落结构的初步研究. 北京师范学院学报, 11(3): 59–64. |
| [] | 陈卫, 高武. 1991. 北京松山自然保护区兽类调查报告. 北京师学院学报:自然科学版, 12(1): 64–68. |
| [] | 陈志平, 王应祥, 冯庆, 等. 1996. 云南西双版纳片断热带雨林鼠形啮齿类的物种多样性研究. 动物学研究, 17(4): 451–458. |
| [] | 王淯, 胡锦矗, 谌利民. 1997. 用最优分割法对啮齿类群落进行分类的探讨. 四川师范学院学报:自然科学版, 18(2): 90–95. |
| [] | 王淯, 王小明, 胡锦矗, 等. 2003. 唐家河自然保护区小型兽类群落结构. 兽类学报, 23(1): 39–44. |
| [] | 吴德林, 罗成昌. 1993. 人类活动对云南哀牢山小型兽类群落结构的影响. 动物学研究, 14(1): 35–41. |
| [] | 武晓东, 施大钊, 刘勇, 等. 1994. 库布其沙漠及其毗邻地区鼠类群落结构分析. 兽类学报, 14(1): 43–50. |
| [] | 武晓东, 邹元平, 郝玉山, 等. 1999. 内蒙古大兴安岭吉文林区啮齿动物区系及危害的调查. 内蒙古农牧学院学报(2): 23–28. |
| [] | 武晓东, 付和平. 2000. 内蒙古半干旱区鼠类群落结构及鼠害危害类型的研究. 兽类学报, 20(1): 21–29. |
| [] | 武晓东, 阿娟, 付和平. 2003. 内蒙古阿拉善荒漠啮齿动物群落结构及其多样性研究. 草地学报, 11(4): 312–316. DOI:10.11733/j.issn.1007-0435.2003.04.007 |
| [] | 袁帅, 武晓东, 付和平, 等. 2011. 不同干扰下荒漠啮齿动物群落多样性的多尺度分析. 生态学报, 31(7): 1982–1992. |
| [] | 杨扬, 朱星, 赵豫桥, 等. 2008. 北京小龙门地区啮齿类群落结构及变化调查. 安徽农业科学, 36(4): 1597–1598. |
| [] | 张洁. 1984. 北京地区鼠类群落结构的研究. 兽类学报, 4(4): 265–271. |
| [] | 张美文, 王凯荣, 王勇, 等. 2003. 洞庭湖区鼠类群落的物种多样性分析. 生态学报, 23(11): 2260–2270. DOI:10.3321/j.issn:1000-0933.2003.11.008 |
| [] | 张知彬, 王福生. 2001. 鼠类对山杏种子存活和萌发的影响. 生态学报, 21(11): 1761–1768. DOI:10.3321/j.issn:1000-0933.2001.11.003 |
| [] | 周庆强, 钟文勤, 孙崇潞. 1982. 内蒙古白音锡勒典型草原区鼠类群落多样性的研究. 兽类学报, 2(1): 89–94. |
| [] | 周旭东, 张永军, 黄健, 等. 2004. 新疆甘家湖自然保护区啮齿动物群落结构与时间动态分析. 动物学杂志, 39(6): 58–61. |
| [] | 曾宗永. 1994. 北美CHIHUAHUAN荒漠啮齿动物群落动态Ⅰ. 年间变动和趋势.兽类学报, 14(1): 24–34. |
| [] | Aragon E E, Garza A, Cervantes F A. 2009. Structure and organization of rodent assembles of a forest of the Sierra Madre Occidental, Durango, Mexico. Revista Chilena de Historia Natural, 82(4): 523–542. |
| [] | Courtalon P, Busch M. 2010. Community structure and diversity of sigmodontine rodents (Muridae: Sigmodontinae) inhabiting maize and soybean field in pampean agroecosystems, Argentina. Interciencia, 35(11): 812–817. |
| [] | Grant W E, Bireny E C. 1979. Small mammal community structure in North American Grasslands. J Mamm, 60(1): 23–36. DOI:10.2307/1379755 |
| [] | Margalef D R. 1958. Information theory in ecology. General System, 3: 36–71. |
| [] | Mangan S A, Ader G H. 2000. Consumption of arbuscular mycorrhizal by terrestial and arboreal small mammals in Panamanian cloud forest. J Mamm, 81(2): 563–570. DOI:10.1644/1545-1542(2000)081<0563:COAMFB>2.0.CO;2 |
| [] | Pielou E C. 1966. The measurement of diversity in different types of biological collection. Journal of Theoretical Biology, 13: 131–144. DOI:10.1016/0022-5193(66)90013-0 |
| [] | Simpson E H. 1949. Measurement of diversity. Nature, 163: 688. DOI:10.1038/163688a0 |
| [] | Tibault K M, Ernest S K M, White E P, et al. 2010. Long-term insights into the influence of precipitation on community dynamics in desert rodents. Journal of Mammalogy, 91(4): 787–797. DOI:10.1644/09-MAMM-S-142.1 |
| [] | Whittaker R H. 1960. Vegetation of the Siskiyou Mountains Oregon and California. Ecological Monographs, 30(3): 279–338. DOI:10.2307/1943563 |