 2012, Vol. 48
2012, Vol. 48文章信息
- 闫守刚, 许清涛
- Yan Shougang, Xu Qingtao
- 流动沙丘与丘间低地过渡带上芦苇幼苗的出土和定居*
- Seedling Emergence and Establishment of Phragmites communis in Transition Zones between Active Dunes and Interdune Lowlands of Active Dune Fields
- 林业科学, 2012, 48(9): 50-55.
- Scientia Silvae Sinicae, 2012, 48(9): 50-55.
-
文章历史
- 收稿日期:2011-10-27
- 修回日期:2012-02-09
-
作者相关文章



流动沙丘区植被恢复常始于丘间低地植被自然恢复,而沙丘多年生植物在流动沙丘与丘间低地过渡带的幼苗出土和定居是丘间低地植被恢复的起源(曹成有等,2000)。由于沙丘区环境恶劣,风沙活动强烈,沙丘多年生植物演化形成了独特的繁殖策略适应风沙活动(Yu et al.,2002;Zhao et al.,2002; 闫巧玲等,2004; 刘志民等,2005)。
芦苇(Phragmitescommunis)进行营养繁殖和有性繁殖。许多研究表明:芦苇种群扩展主要依靠营养繁殖,因为芦苇实生幼苗很少被发现,尽管芦苇每年都开花、结种、种子成熟(Grace et al.,1981;1982; Bartley et al.,1987)。然而,野外观察表明:芦苇有性繁殖苗在生长季初期出现在风蚀作用强烈和土壤水分较好的流动沙丘与丘间低地过渡带上。因此假设:导致良好土壤水分的风蚀作用,有利于芦苇的种子萌发、出苗和定居。为了检验这一假设,通过野外调查,监测了流动沙丘与丘间低地过渡带的风蚀深度、地下水位、土壤水分以及芦苇的土壤种子库、实生幼苗。
与其他物种相比,芦苇有较早的有性繁殖和较长的生长期,这使得其生境向着适宜植被生长的方向发展。因此假设:芦苇的有性繁殖促进了丘间低地植被的自然恢复,在流动沙丘区的植被恢复中也起重要作用。为了检验这一假设,在植物生长季末调查了芦苇实生幼苗的定居情况。
1 研究方法 1.1 样地选择与设置研究区位于科尔沁沙地西部乌兰敖都地区。当地年最高和最低气温分别出现在8月和1月,生长季(5—9月)月均温在16.3~22.2℃。该区域属于温带大陆性半干旱气候,年均气温6.2℃,年均降水量340.5mm,多集中在7—8月。风沙大且频繁,年均风速为3.5~4.5m·s-1。3—5月为风季,盛行偏西北或偏北风。地带性土壤主要为沙质栗钙土,被破坏后则退化为流动风沙土。
样地位于流动沙丘区中心,沙丘高15~25m,每年以5~7m速度前移,植被盖度低于5%,是当地典型流动沙丘(蒋德明等,2003)。沙丘先锋植物沙蓬(Agriophyllum squarrosum)和乌丹蒿(Artemisia wudanica)分布较广,群落结构单一,常形成单优群落。包围在新月形沙丘中的丘间低地,具有良好水分条件,群落类型多样,种类丰富,并发育大量非沙生植物,如小香蒲(Typhaminima)、拂子茅(Calamagrostis epigeios)等。
在2个沙丘区,各选择1块典型丘间低地,调查丘间低地面积、土壤和植被状况,确定流动沙丘与丘间低地之间过渡带的位置,并按方位顺序编号为过渡带1和2。
1.2 研究方法2009年11月,在每个过渡带设置4条样带,每条样带间隔1 m设置7根1 m的铁签。过渡带1和2分别设置28个铁签。从2010年4月初到9月末每10天测量一次铁签的高度,监测过渡带风蚀深度的动态变化。
2010年4月初,在每个过渡带设置4条样带,每条样带间隔1 m设置7个1 m × 1 m的样方对。过渡带1和2分别设置28个样方对。一个样方用来监测芦苇实生幼苗的出土动态(称为出苗样方),另一个样方用来监测生长季末芦苇幼苗的定居数量(称为定居样方)。
在2010年5月初到9月底,在每个出苗样方调查: 0 ~ 30 cm(3层: 0 ~ 5,5 ~ 15,15 ~ 30 cm)土壤水分、地下水位、0 ~ 5 cm土壤种子库、实生幼苗。生长季末,在定居样方调查芦苇幼苗的定居情况。
1) 土壤水分监测方法 从2010年5月初,每10天调查每个出苗样方0 ~ 30 cm(3层: 0 ~ 5 cm,5 ~ 15 cm,15 ~ 30 cm)深度土壤含水量。采用烘干称量法测定土壤含水量,即用直径5 cm的土钻在每个样方附近取土样(体积98 cm3),用百分之一天平称鲜质量,在烘箱(105 ℃)中烘8 h,再称量。
2) 地下水位监测方法 在每个过渡带上打一口井,将一个塑料管永久性地放入井中。从2010年5月初,每10天量测地下水位。
3) 土壤种子库监测方法 从2010年5月初开始,在每个样方附近,每10天取一次0 ~ 5 cm的土壤种子库。直径7 cm的环刀取土样(Yan et al.,2007),土样风干后采用物理分离法确定土壤种子库大小,即用粒径0. 5 mm的筛子筛选,并把土样中的芦苇种子手工挑出,最后用TTC法检验种子的活力(Paker et al.,1996)。
4) 芦苇幼苗出土监测方法 从2010年5月初,每10天监测、计数并拔出每个出苗样方内出土的有性繁殖苗。
5) 芦苇幼苗定居的调查方法 在2010年9月底,记录每个定居样方内芦苇的实生苗的定居植被数量和高度。
1.3 数据分析方法用SPSS 11. 5软件分析芦苇的实生幼苗与不同深度土壤水分的相关性,P<0. 05被认为相关性显著。
2 结果与分析 2.1 芦苇土壤种子库在野外调查过程中,在过渡带0 ~ 5 cm的土壤样品中很少发现有活力的种子(≤3粒·m-2),因此土壤种子库的数据未被列出。
2.2 芦苇实生幼苗出土的时间格局流动沙丘与丘间低地过渡带1和2的芦苇幼苗出土呈单峰曲线,幼苗在5月18日开始出土,5月28日达到出苗高峰,80. 4%的幼苗在5月18日到6月18日的1个月内出土(图 1)。

|
图 1 芦苇幼苗出土的时间格局(平均值±标准误) Fig.1 Temporal patterns of seedling emergence (M ± SE) of P. communis |
过渡带1的芦苇幼苗出土最大密度为(13.5± 1.22)株·m-2,出现在5月28日; 过渡带2的芦苇出土幼苗密度有2个高峰值,为(6.3±1.18)株·m-2和(6.5±0.83)株·m-2,分别出现在5月28日和6月8日。从6月28日到9月8日,在每个调查时间点上,芦苇出土幼苗密度都<2株·m-2(图 1)。
2.3 芦苇实生幼苗与土壤水分的关系芦苇出土幼苗密度与表层土壤水分的相关性随幼苗出土时间的不同而不同。在出苗高峰期,出土幼苗数量与土壤水分呈显著正相关(P<0. 05) (表 1),在其他时间段,二者的相关性未达到显著水平(P>0. 05)。
|
|
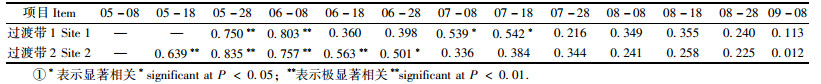
在幼苗出土高峰期(5月28日),芦苇幼苗密度与土壤水分呈对数关系(图 2)。随着土壤水分的增加,样方内的幼苗密度呈对数增加。对于过渡带1来说,土壤水分>3. 0%时芦苇幼苗才会出土,对于过渡带2来说,土壤水分>3. 7%时芦苇幼苗才会出土,即芦苇实生幼苗出土的土壤水分阈值分别为3. 0%和3. 7%。

|
图 2 芦苇实生幼苗密度和土壤水分的对数函数关系(a.过渡带1;b.过渡带2) Fig.2 Relationships in seedling density of P. communis and soil moisture of quadrats at site 1 (a) and site 2 (b) |
总体来看,从丘间低地迎风坡边缘到流动沙丘迎风坡坡脚,过渡带1和2的芦苇幼苗出土个数与表层土壤水分呈显著正相关(P<0. 05) (图 3)。

|
图 3 芦苇实生苗密度与土壤水分的相关性(a,过渡带1; b,过渡带2) Fig.3 Correlation in seedling density and soil moisture of quadrats at site 1 (a) and site 2 (b) 从样方1到样方7的方向是沿着样带从丘间低地迎风坡边缘到流动沙丘迎风坡坡脚 The direction from quadrat 1 to 7 is along the transect from the border of windward slope of the interdune lowland to the foot of the windward slope of the active dune |
过渡带1和2的实生幼苗出土呈现出明显不同的格局。在过渡带1,沿着样带从丘间低地迎风坡边缘到流动沙丘迎风坡坡脚,芦苇幼苗密度先增加,后减少,在土壤水分最好的样方2内,芦苇幼苗密度最大; 在过渡带2,沿着样带从丘间低地迎风坡边缘到流动沙丘迎风坡坡脚,芦苇幼苗密度逐渐减少,样方1的密度最大(图 3)。
2.5 芦苇实生幼苗的定居流动沙丘与丘间低地过渡带1和2出土芦苇幼苗的54. 1%在生长季末定居; 定居植被密度与出土幼苗密度呈显著正相关(P<0. 05) (图 4)。

|
图 4 芦苇出土幼苗数与生长季末定居植物个数的相关性(a,过渡带1; b,过渡带2) Fig.4 Seedling emergence and seedling establishment in different quadrats at site 1 (a) and site 2 (b) “幼苗密度”指每隔10天出苗样方出土的芦苇幼苗总数,“植物密度”指生长季末定居样方定居的芦苇数量。从样方1到样方7方向是沿样带从丘间低地迎风坡边缘到流动沙丘迎风坡坡脚。 "Seedling density"means the seedling density of P. communis observed at 10 d intervals in each emergence quadrat, and"plant density"implies the plant density of P. communis established in each establishment quadrat throughout a whole growing season. The direction from quadrat 1 to quadrat 7 is along the transect from the border of windward slope of the interdune lowland to the foot of the windward slope of the active dune |
植物种群分布与种子传播、萌发及幼苗出土、定居密切相关(Sagrario et al.,2005)。Whipple(1978)将地上植被和土壤种子库之间的关系分为4种类型:类型Ⅰ,种子和地上植被都存在; 类型Ⅱ,只有种子没有植被; 类型Ⅲ,只有植被,没有种子; 类型Ⅳ,种子和植被都不存在。芦苇属于类型Ⅲ,有植被,但土壤种子库中没有种子。
芦苇种子单粒质量(0. 32 ± 0. 01) mg(闫巧玲等,2004),种子小且具有冠毛的特征体现了其有性繁殖的内在特性。根据观察,芦苇种子在每年的11月成熟,种子传播出现在第2年的风季,大风促进了种子的传播。另外,芦苇种子一接触到湿润的地表就快速萌发。
丘间低地外部条件(即风蚀和水文状况)对于芦苇幼苗出土和定居至关重要。随着流动沙丘前移,在沙丘后部就会出现一个空白带,即流动沙丘与丘间低地过渡带(曹成有等,2000)。过渡带上风蚀很强烈,深度达到32 ~ 35 cm,使地表更接近地下水位(24 ~ 28 cm),0 ~ 5 cm土壤水分﹥ 9%。这样,良好的土壤水分更利于捕获种子和种子的萌发,这是因为芦苇种子既小又具冠毛,更易黏附于湿润的地表,从而促进了芦苇种子的萌发过程。在野外,只有当土壤水分达到3. 0%的时候,才可能出现芦苇幼苗(图 2)。可见,风蚀是促进芦苇种子萌发、幼苗出土和幼苗定居的一个重要驱动力。
良好的水文状况是流动沙丘区芦苇种子萌发、幼苗出土和定居的重要保证。目前的研究表明芦苇幼苗出土更多依赖于土壤水分(表 1,图 3),而土壤水分更多依赖于地下水的供给(Yan et al.,2007)。在丘间低地,地下水通过毛细管作用不断向地面移动,这样就维持了土壤水分。根据野外观察,虽然在芦苇母株下有很多芦苇种子,但是却很少有其幼苗。芦苇母株的生长会降低土壤水分(刘慎谔文集编委会,1985),因此土壤水分不能满足芦苇种子在母株下萌发。可见,土壤水分是限制芦苇种子萌发、幼苗出土和定居的主要因子。同时也说明,芦苇成年母株的生长,不利于种子在其周围进行有性繁殖,进而不利于其在流动沙丘区的植被恢复。
综上所述,得出以下结论: 1)芦苇种子在每年11月成熟,在第2年风季传播; 2)芦苇种子一接触到湿润的地表就萌发,且种子萌发更多依赖于土壤水分。芦苇种子一落到地表,要么快速萌发,要么就被风吹走,这就是为什么在丘间低地中芦苇土壤种子库非常少的原因。
3.2 芦苇在过渡带植被恢复中的作用在一个风蚀强烈(深度32 ~ 35 cm)的过渡带,地下水位升高(24 ~ 28 cm)导致土壤水分良好(>9%) (Yan et al.,2007)。同时,在风的作用下,芦苇的成熟种子四处传播。带冠毛的种子黏附在过渡带湿润的土壤表层,从而在此停留、聚集,在温度适宜的时候萌发。出土幼苗发展成为成年植株,形成一处新的芦苇种群斑块,在过渡带裸沙区开创了一个新的植被演替阶段,从而在流动沙丘区植被自然恢复过程中扮演重要角色。
3.3 多年生植物在流动沙丘区植被恢复中的作用多年生植物利用独特的繁殖策略适应风蚀和沙埋,在沙丘植被自然恢复过程中起重要作用(朱震达,1963)。除芦苇外,乌丹蒿(Artemisia wudanica)以独特的繁殖方式(植冠种子库、持久土壤种子库和黏液繁殖体),利用种子进行有性繁殖(闫巧玲等,2004;刘志民等,2005; Yan et al.,2007)。此外,黄柳(Salix gordejevii)在流动沙丘与丘间低地过渡带利用风传播的种子进行有性繁殖(Yan et al.,2007)。
流动沙丘区植被的自然恢复需要通过2种途径来实现:流动沙丘的植被恢复和丘间低地的植被恢复,目前更多的注意力集中在流动沙丘的植被恢复上(Francoise et al.,2004)。然而,丘间低地植被恢复在沙丘区的植被恢复中也起重要作用,因为沙丘区植被自然恢复多数情况下开始于丘间低地植被恢复。流动沙丘前移对丘间低地植被过程的影响主要有2个方面:背风坡沙子前移掩埋丘间低地植被,过渡带风蚀导致新生植被在此定居和扩展。二者的消长调节丘间低地植被的面积和格局,进而调节整个流动沙丘区植被的面积和格局。沙丘多年生植物通过有性繁殖在过渡带定居种群开创了一个新的植被演替过程,扩大了丘间低地植被的面积,形成了不同植被格局(Vitousek et al.,1989)。
总之,流动沙丘和丘间低地之间过渡带上多年生植物幼苗的出土和定居在流动沙丘区植被恢复方面发挥重要作用。
| [] | 曹成有, 寇振武, 蒋德明, 等. 2000. 科尔沁沙地丘间地植被演变的研究. 植物生态学报, 24(3): 262–267. |
| [] | 蒋德明, 刘志民, 曹成有, 等. 2003. 科尔沁沙地荒漠化过程与生态恢复. 北京, 中国环境科学出版社. |
| [] | 刘慎谔文集编委会. 1985. 刘慎谔文集. 北京, 科学出版社. |
| [] | 刘志民, 阎巧玲, 骆永明, 等. 2005. 不同降水量处理时5种蒿属植物瘦果粘液溶出. 生态学报, 25(6): 1497–1501. |
| [] | 闫巧玲, 刘志民, 骆永明, 等. 2004. 科尔沁沙地78种植物繁殖体重量和形状比较. 生态学报, 24(11): 2422–2429. DOI:10.3321/j.issn:1000-0933.2004.11.011 |
| [] | 朱震达. 1963. 风力作用下沙丘演化的动力过程研究. 地理学论文集(第5集). 北京: 中国科学出版社. |
| [] | Bartley M R, Spence D H N. 1987. Dormancy and propagation in helophytes and hydrophytes. Arch Hydrobiol Beih, Ergebnisse der Limnologie, 27(2): 139–155. |
| [] | Francoise R, Servane L. 2004. Sand dune restoration in North Brittany, France: a 10-year monitoring study. Restoration Ecology, 12(1): 29–35. DOI:10.1111/rec.2004.12.issue-1 |
| [] | Grace J B, Wetzel R G. 1981. Effects of size and growth rate on vegetative reproduction in Typha. Oecologia, 50(3): 158–161. |
| [] | Grace J B, Wetzel R G. 1982. Variations in growth and reproduction within populations of two rhizomatous plant species: Typha latifolia and Typha angustifolia. Oecologia, 53(5): 258–263. |
| [] | Paker C E, Venable D L. 1996. Seed bank in desert annuals:implications for persistence and coexistence in variable environments. Ecology, 77(2): 1427–1435. |
| [] | Sagrario A, Francisco S. 2005. Spatial dynamics of Ilex aquifolium populations seed dispersal and seed bank: Understanding the first steps of regeneration. Plant Ecology, 177(3): 237–248. |
| [] | Vitousek P M, Walker L R. 1989. .Biological invasion by Myrica faya in Hawaii: plant demography, nitrogen fixation, and ecosystem effects. Ecological Monographs, 59(3): 247–256. DOI:10.2307/1942601 |
| [] | Whipple S A. 1978. The relationship of buried, germinating seeds to vegetation in an old-growth Colorado subalpine forest. Canadian Journal of Botany, 56(3): 1506–1509. |
| [] | Yan Q L, Liu Z M, Ma J L, et al. 2007. The role of reproductive phenology, seedling emergence and establishment of perennial Salix gordejevii in active sand dune fields. Annals of Botany, 99(1): 19–28. DOI:10.1093/aob/mcl228 |
| [] | Yu F H, Chen Y F, Dong M. 2002. Clonal integration enhances survival and performance of Potentilla anserina, suffering from partial sand burial on Ordos plateau, China. Evolutionary Ecology, 15(4): 303–318. |
| [] | Zhao W Z, Liu Z M. 2002. Responses of growth and reproduction of Sophora moorcroftiana to altitude and sand-burying in Tibet. Acta Ecologica Sinica, 22(1): 134–138. |