 2012, Vol. 48
2012, Vol. 48文章信息
- 赵猛, 魏朔南, 胡正海
- Zhao Meng, Wei Shuonan, Hu Zhenghai
- 漆树韧皮部的结构与发育*
- Structure and Development of Phloem in Toxicodendron vernicifluum
- 林业科学, 2012, 48(9): 36-41.
- Scientia Silvae Sinicae, 2012, 48(9): 36-41.
-
文章历史
- 收稿日期:2012-01-04
- 修回日期:2012-06-24
-
作者相关文章



2. 山西师范大学 临汾 041004
2. Shanxi Normal University Linfen 041004
漆树(Toxicodendron vernicifluum)为漆树科(Anacardiaceae)漆树属(Toxicodendron)落叶乔木,主要分布在我国陕西、湖北、四川、重庆、甘肃、贵州和云南等省(市) (张飞龙等,2007),在泰国、越南、日本以及朝鲜半岛等地也有分布(Beaman,1986; Hong et al.,1999)。漆树的次生代谢产物生漆是一种优良的天然涂料,漆树的栽培与生漆的应用在我国及东亚、东南亚各国有着悠久的历史(Vogl,2000; Webb,2000; 张飞龙,2006)。漆树的韧皮部是生漆产生和贮存的主要部位,生漆的采集通常是在漆树主干树皮上割口,使生漆流出并收集而获得(汪明正等,2008; 胡正海等,1979)。20世纪70年代以后,在漆树的种质资源及其分布(张飞龙等,2007)、乳汁道及其周围组织结构(胡正海等,1979; 赵桂仿等,1990; 1991)以及生漆化学( Hong et al.,1999; Kim et al.,2009; Yamauchi et al.,1982)等方面已有许多研究报道。但是,对漆树的经济利用部分茎干韧皮部的结构及其发育规律尚缺乏系统的研究报道。
树木次生韧皮部发育的季节性变化的研究可为了解植物生长发育规律提供资料,并能探讨外界环境因素对植物生长发育的影响(张振珏,1991),而且韧皮部发育与植物次生代谢产物合成的关系也是目前植物结构与功能研究中备受关注的问题(Samanani et al.,2006)。自20世纪50年代以来,已有北美乔松(Pinus strobus) (Alfieri et al.,1973)、卡拉里亚松(Pinus brutia) (Ewers et al.,1987)、复叶槭(Acer negundo) (Tucker et al.,1969)、欧洲云杉(Picea abies) (Wimmer et al.,1997)等多种热带及温带树木进行了韧皮部的结构和发育的研究; 我国学者也对华北地区的多种温带树木进行了相关的韧皮部发育研究(张英伯等,1982),另有学者对产于我国亚热带地区的黄檀属(Dalbergia)及多种单宁植物的韧皮部发育进行了研究(张振珏等,1994a; 1997),并探讨了女贞子(Ligustrum lucidum)以及白蜡树( Fraxinus chinensis)韧皮部的季节性变化,为药用部分的结构、生长和发育提供了资料(张振珏等,1994b)。为此,树木韧皮部结构与发育的研究具有一定理论和实践意义。
本文以我国生漆主产区陕西省的3个漆树品种的主干韧皮部为材料,研究其结构特点,并系统采集不同月份的韧皮部材料,研究韧皮部结构的发育过程和季节性变化,阐明漆树韧皮部的结构和发育规律,为改进生漆的采割技术提供理论依据。并为植物次生韧皮部组成分子的发育与其次生代谢产物的合成与分泌的关系研究提供理论依据。
1 材料与方法 1.1 试验材料陕西省是我国生漆的主要产区,以引种栽培于陕西省西安植物园的2个栽培漆树品种‘大红袍’ (T. vernicifluum ‘Dahongpao’)、‘高八尺’(T. vernicifluum‘Gaobachi’)和1个野生漆树品种‘宁陕’(T. vernicifluum ‘Ningshan’)各3株为研究材料,树龄均为15年,凭证标本保存于西北大学生命科学学院植物标本馆。为研究漆树次生韧皮部各组成分子的发育和季节性变化,2010年全年逐月在漆树主干高约1 ~ 2 m处采集3 cm × 3 cm韧皮部材料1块,采用FAA溶液固定(甲醛:冰醋酸: 70%酒精= 5: 5: 90,V /V); 冰冻切片所需材料带回实验室恒温冷冻保存,保存温度- 15 ℃。
1.2 研究方法 1.2.1 石蜡切片材料经FAA溶液固定1周后用软化剂(70%乙醇:甘油= 1: 1,V /V)软化60天,室温下依次采用70%,80%,90%,100%酒精梯度脱水(每一梯度30 min),二甲苯透明,渗蜡,石蜡包埋,Leica RM2135切片机切片,切片厚度6 ~ 8 μm,番红–固绿染色,中性树胶封片; Nikon Eclipse 50i显微镜观察,DS-Fi1成像系统拍照。
1.2.2 半薄切片将试验材料分割为合适大小,戊二醛(4%)、锇酸(1%)固定,酒精脱水,环氧丙烷过渡,Epon 812树脂包埋,Reichert-Jung超薄切片机切片,切片厚度1 ~ 2 μm,甲苯胺蓝染色。Nikon Eclipse 50i显微镜观察,DS-Fi1成像系统拍照。
1.2.3 冰冻切片及漆酚类物质组织化学检测将试验材料分割为合适大小,Tissue Freezing Medium包埋,Leica CM1850冰冻切片机切片,切片厚度25 ~ 30 μm,三氯化铁–乙醇溶液检测漆酚类物质的存在。
1.2.4 木质素组织化学试验石蜡制片脱蜡后,滴加间苯三酚乙醇溶液和浓盐酸,对漆树韧皮部石细胞中所含的木质素进行组织化学检测,Nikon Eclipse 50i显微镜观察,DS-Fi1成像系统拍照。
2 结果与分析 2.1 漆树韧皮部的结构特征通过3个漆树品种韧皮部的各个切面的显微结构观察后发现,不同品种间漆树韧皮部的基本结构特征相似,其内、外侧分别分布有维管形成层和周皮(图版Ⅰ-1 ~ 3)。根据发育时期和组织结构特点的差异,次生韧皮部在横切面上又可分为具功能韧皮部和无功能韧皮部2部分。具功能韧皮部与维管形成层相连,为当年产生和分化形成的次生韧皮部,由筛管、伴胞、韧皮薄壁细胞和尚处于形成阶段的乳汁道组成(图版Ⅱ– 1),上述组成分子被单列或多列的韧皮射线细胞所分隔。每年早期形成的韧皮部的生长层可以辨认出来,此部分韧皮部中筛管细胞饱满、明显,薄壁组织细胞狭小,称为具功能韧皮部。无功能韧皮部位于具功能韧皮部外侧,为往年形成的次生韧皮部,无功能韧皮部的外侧与周皮相连。无功能韧皮部的主要结构特点为筛管萎缩变形,失去输导功能,韧皮薄壁细胞的体积增大,乳汁道已发育成熟(图版Ⅱ– 2);同时,无功能韧皮部中分化出石细胞(图版Ⅰ-1 ~ 3),而且韧皮射线细胞由单列发育成2 ~ 4列。无功能韧皮部外侧为周皮,由木栓层、木栓形成层和栓内层组成(图版Ⅱ– 3)。

|
图 图版Ⅰ 1.‘大红袍’漆树韧皮部横切面; 2.‘高八尺’漆树韧皮部横切面; 3.‘宁陕’漆树韧皮部横切面。上述材料均采自2010年8月生漆采割季节。 Fig.图版Ⅰ 1. Phloem cross section of'Dahongpao'; 2. Phloem cross section of 'Gaobachi'; 3. Phloem cross section of'Ningshan'. These materials were collected in August 2010. C:形成层Cambium; FP:具功能韧皮部Functional phloem; PEL:木栓形成层Phellogen; PHE:木栓层Phellem; PR:韧皮射线Phloem rays; SC:石细胞Stone cells. |

|
图 图版Ⅱ 1.具功能韧皮部横切面,示各组成分子; 2.无功能韧皮部横切面,示各组成分子及发育成熟乳汁道; 3.周皮横切面,示木栓层、木栓形成层和栓内层细胞; 4.具功能韧皮部横切面,示韧皮薄壁细胞与筛管分子带状排列; 5.具功能韧皮部纵切面,示韧皮薄壁细胞与筛管分子带状排列; 6.发育期狭缝状乳汁道横切面,示分泌细胞与鞘细胞; 7.成熟乳汁道横切面,示分泌细胞与鞘细胞; 8.形成层及具功能韧皮部横切面,示多列射线细胞; 9.具功能韧皮部与无功能韧皮部横切面,示二者界限。 Fig.图版Ⅱ 1. Cross section of functional phloem, show all composed of molecules of fun-phloem; 2. Cross section of non-functional phloem, show all composed of molecules and mature laticiferous canals; 3. Cross section of periderm, show pellem, phelloderm and phellogen; 4. Cross section of functional phloem, show the parenchyma cells and sieve tube arranged in ribbon; 5. Longitudinal section of functional phloem, show the parenchyma cells and sieve tube arranged in ribbon; 6. Laticiferous canals in the developmental stages, show secreting cells and sheath cells; 7. Cross section of mature laticiferous canals, show secreting cells and sheath cells; 8. Cross section of functional phloem and cambium, show phloem ray; 9. Cross section of functional phloem and non-functional phloem, show the border between them. 材料Materia:l ‘高八尺’漆树Toxicodendron vernicifluum‘Gaobachi’; 采集日期Collecting date: 1,4,5,6. 2010-05; 2,3. 2010-07; 7,8,9. 2010-08. C:形成层Cambium; CC:伴胞Companion cell; FP:具功能韧皮部Functional phloem; NP:无功能韧皮部Non-functional phloem; PE:栓内层Phelloderm; PEL:木栓形成层Phellogen; PHE:木栓层Phellem; PP:韧皮薄壁细胞Phloem parenchyma; PR:韧皮射线Phloem rays; SE:上皮细胞Epithelia; SH:鞘细胞Sheath; ST:筛管分子Sieve tubes; ★:漆树乳汁道Laticiferous canals of lacquer tree. |
3个漆树品种的韧皮部都具有上述特征,但品种间的韧皮部总厚度存在显著差异(表 1)。
|
|
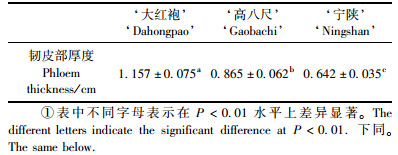
漆树的茎干树皮与绝大多数裸子植物和被子植物中的双子叶植物一样,是由其维管形成层细胞发生平周分裂,在一定条件下由其靠外的衍生细胞分化而成,其发育过程可归纳如下。
2.2.1 具功能韧皮部的分化和发育漆树茎干维管形成层向外产生的衍生细胞逐步分化为具功能韧皮部,在垂直系统中,筛管和伴胞最早分化成熟,在当年产生的次生韧皮部的横切面上,可见临近维管形成层区处筛管发育为近多角形,细胞形态饱满; 筛管旁侧的小型细胞为伴胞,筛管和伴胞呈带状排列,占据具功能韧皮部大部分区域(图版Ⅱ– 4)。在弦向纵切面上,筛管分子呈纺锤形,薄壁细胞体积较小(图版Ⅱ– 5)。在筛管分化发育同时,韧皮薄壁细胞也开始分化; 韧皮薄壁细胞分布在筛管之间,也呈带状排列,当年产生的韧皮薄壁细胞其体积尚未增大(图版Ⅱ– 7,8)。乳汁道的分化早期是由多个呈莲座状排列的原始细胞组成,以后它们间的中层溶解,裂生成狭缝状腔(图版Ⅱ– 6),随后乳汁道的腔隙逐渐扩大,可见多角形等多种形状,最终在横切面上形成圆形的腔隙,乳汁道腔周围被1层分泌细胞和2 ~ 3层鞘细胞包围(图版Ⅱ– 7)。
具功能韧皮部径向系统的韧皮射线由射线原始细胞的衍生细胞形成,与垂直系统的组成分子同时分化,当年产生的韧皮射线通常为单列或双列细胞,偶见3 ~ 5列,其细胞质浓厚(图版Ⅱ– 8),在次生韧皮部中由内向外呈辐射状排列,从而将垂直系统组成分子隔成条块状(图版Ⅰ– 1 ~ 3)。在当年产生的具功能韧皮部中,筛管和伴胞已经发育成熟,韧皮薄壁细胞已分化,但尚未发育成熟; 乳汁道处于分化、发育过程中,以上组成分子被韧皮射线辐射分割。由于此部分次生韧皮部中筛管具有输导功能,为此,通常将当年由维管形成层产生的次生韧皮部称为具功能韧皮部。所观察的3个漆树品种的具功能韧皮部结构相似,但品种间具功能韧皮部的厚度差异显著(表 2)。
|
|
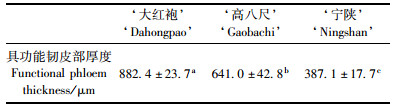
无功能韧皮部是指当年由维管形成层向外产生的具功能韧皮部以外的次生韧皮部的总称,通常占据茎干次生韧皮部的大部分区域(图版Ⅰ– 1 ~ 3)。无功能韧皮部的筛管已失去输导功能,其他组成分子的结构也发生相应变化,并产生新的组织。从横切面观察,无功能韧皮部与具功能韧皮部之间在结构上显示出明显界限(图版Ⅱ– 9),在无功能韧皮部中,各类组成分子由内向外发生明显的结构变化,从横切面上观察,韧皮薄壁细胞体积进一步增大并挤压筛管使其萎缩、变形,细胞截面减小,形状不规则(图版Ⅲ– 1)。从弦向纵切面上观察,薄壁细胞体积增大,筛管受到挤压而显细长(图版Ⅲ– 2,与图版Ⅱ– 5比较)。乳汁道散生在2列射线之间,排列成单列,少数为2列,乳汁道之间由其他组成分子所分隔(图版Ⅰ– 1 ~ 3)。在具功能韧皮部中,漆树乳汁道处于发育初期,而在相邻的无功能韧皮部中乳汁道已发育成熟,由2 ~ 3层鞘细胞、1层分泌细胞包围着近圆形腔道构成,乳汁道腔内充满生漆(图版Ⅲ– 3)。邻近具功能韧皮部的无功能韧皮部中的乳汁道是产生和贮存生漆的主要场所,而在邻近周皮的乳汁道中,由于其茎干中形成层的连续活动,木质部大量增加而直径增大,而外面周皮周长的增加滞后且少,故将乳汁道腔挤压变为切向椭圆形,其分泌细胞萎缩,失去产生和分泌生漆的功能(图版Ⅲ– 4);同时还观察到部分乳汁道中产生拟侵填体,将乳汁道腔堵塞(图版Ⅲ– 5)。

|
图 图版Ⅲ 1.无功能韧皮部横切面,示次生韧皮部中筛管萎缩变形,韧皮薄壁细胞的体积增大,数量增多; 2.无功能韧皮部纵切面,次生韧皮部中筛管呈长形; 3.乳汁道漆酚组织化学反应; 4.近周皮处无功能韧皮部,示乳汁道切线椭圆形; 5.乳汁道腔中拟侵填体细胞; 6.无功能韧皮部中木质素组织化学检测,示石细胞中木质素沉积位置; 7.韧皮部石细胞次生壁加厚形成石细胞群; 8.韧皮部石细胞次生壁分层加厚; 9.落皮层,示落皮层中残存的石细胞群和乳汁道。 Fig.图版Ⅲ 1. Cross section of non-functional phloem, show the shape of sieve tube shrinked and parenchyma cells increased in size and quantity; 2. Longitudinal section of non-functional phloem, show the shape of sieve tube elongated; 3. Histochemical test of raw lacquer; 4. Nonfunctional phloem which near the periderm, show the form of laticiferous canals change into oval; 5. Tylosoids cells in laticiferous canals; 6. Histochemical test of lignin in non-functional phloem, show the lignin deposition positions; 7. Lignin deposited in secondary wall and formed stone cells; 8. Stone cells, show the deposition of lignin in the cell wall was layered; 9. Periderm, show the stone cells and laticiferous canals remain inside. 材料Material: 1,2. ‘大红袍’漆树Toxicodendron vernicifluum‘Dahongpao’; 3-9: ‘高八尺’漆树T. vernicifluum‘Gaobachi’.采集日期Collecting date: 1,2,3. 2010-08; 4-9. 2010-09. CC:伴胞Companion cell; PP:韧皮薄壁细胞Phloem parenchyma; RL:生漆Raw lacquer; SC:石细胞Stone cells; ST:筛管分子Sieve tubes; TY:拟侵填体Tylosoids; ★:漆树乳汁道Laticiferous canals of lacquer tree. |
在上述组织的发育变化过程中,无功能韧皮部中部分薄壁细胞分化为石细胞群。所观察的3个漆树品种中,无功能韧皮部中均有石细胞群的分布; 从整个无功能韧皮部区域来看,石细胞群在周皮一侧的数量多于临近具功能韧皮部侧,而且面积也较大。从韧皮部横切面观察,石细胞均聚集成簇,面积大小不等(图版Ⅰ– 1 ~ 3)。组织化学研究表明,韧皮薄壁细胞的初生细胞壁内形成次生壁,木质素沉积在次生壁内(图版Ⅲ– 6),往往相邻的许多韧皮薄壁细胞同时发育成石细胞而组成石细胞群(图版Ⅲ– 7),在成熟的石细胞的次生壁中可看到明显的S1,S2和S3层(图版Ⅲ– 8)。
2.2.3 周皮的结构多年生的漆树茎周皮与其他树种一样,由木栓形成层、木栓层和栓内层共同构成(图版Ⅱ– 3)。其木栓形成层可能活动一段时期后就停止活动,由更内部的生活的薄壁细胞重新发生,所以,多年生的漆树中的周皮可能由韧皮薄壁细胞脱分化、再分化形成木栓形成层,木栓形成层向外产生木栓层,向内产生栓内层,共同构成次生保护组织周皮。而且在以后的生长发育中,由于维管形成层的周期性分裂,使漆树茎的直径不断增加,最初产生的周皮连同原来的木栓形成层一起死亡而形成落皮层脱落。落皮层中能看到一些残存的石细胞群和乳汁道(图版Ⅲ– 9),这就间接证明了前述的木栓形成层的发生和活动方式,即后期的木栓形成层是由次生韧皮部的韧皮薄壁组织细胞发生的。
2.2.4 漆树韧皮部结构发育的季节性变化漆树的韧皮部是由茎干的维管形成层细胞向外分裂产生的衍生细胞分化形成的,漆树的维管形成层活动像其他树木一样具有周期性,3个漆树品种中维管形成层活动的强弱也存在差异,从而导致新产生的具功能韧皮部的厚度不同,‘大红袍’>‘高八尺’>‘宁陕’漆树。无功能韧皮部中韧皮薄壁细胞数量的进一步增加以及石细胞群的产生等也多发生在春、夏两季。
乳汁道是漆树产生和储存生漆的结构,无功能韧皮部中的乳汁道已发育成熟,根据对不同季节韧皮部冰冻切片的漆酚类物质组织化学研究表明,每年春末至初秋,乳汁道腔内充满生漆(图版Ⅲ– 3),秋季9月后(割漆季节结束)其腔道中不含或仅含少量生漆; 此外,3个品种漆树无功能韧皮部中在割漆季节结束以后都有部分乳汁道腔内由上皮细胞增殖产生拟侵填体,堵塞腔道(图版Ⅲ– 5)。比较观察3个漆树品种拟侵填体的发生规律,其发生过程相似,但发生时间存在差异,‘大红袍’为9月上、中旬,而‘高八尺’和‘宁陕’漆树为8月中、下旬,拟侵填体的解体时间则一致,都在次年春季。
3 讨论 3.1 漆树韧皮部的结构特征及其研究意义漆树乳汁道在其根、茎、叶、花和果实中均有分布,主要分布于各器官的韧皮部中(傅淑颖等,2007),生漆通常是在割漆季节从漆树主干树皮上割口,使生漆流出后收集而获得。因此,阐明漆树韧皮部的结构特征,可以为改进割漆技术,提高生漆产量提供理论依据,对漆树资源的保护也有重要意义。橡胶植物巴西橡胶树(Hevea brasiliensis)也是从其主干树皮上割口使乳胶液流出而获得。橡胶的树皮从内到外可分为5层,即维管形成层、水囊皮、黄皮、砂皮以及粗皮5部分; 水囊皮为具功能韧皮部,由筛管、伴胞、韧皮薄壁细胞、乳汁管和韧皮射线构成,水囊皮中的乳汁管尚处于发育中; 黄皮为无功能韧皮部的内侧部分,与水囊皮具有相同的组成分子,其中乳汁管已发育成熟,富含橡胶,黄皮中有少量石细胞群出现; 砂皮为无功能韧皮部的中部以外区域,其石细胞群数量增多,乳汁管逐渐衰老,失去产胶能力; 粗皮即为橡胶树的周皮,根据橡胶树皮的上述结构特点,总结提出割胶时要保护水囊皮即具功能韧皮部(郝秉中等,1980)。巴西橡胶树的产胶结构为乳汁管,是1列相互贯通的细胞组成有节乳汁管,其乳胶包括全部细胞质(Hagel et al.,2008);而漆树产生生漆的结构为分泌道,其乳汁是乳汁道分泌细胞产生后分泌到腔道中贮存的次生代谢产物漆酚类物质(胡正海等,1979)。漆树与橡胶树的树皮结构特点相似,漆树的树皮从内到外依照橡胶树皮结构的划分和漆树树皮的发育情况也可分为5层,漆树具功能韧皮部中筛管发育成熟,而乳汁道处于发育初期; 具功能韧皮部外侧的无功能韧皮部类似橡胶树的黄皮区域,乳汁道发育成熟,分泌活动旺盛,而且在这一区域筛管萎缩,失去输导功能,石细胞群少见; 其外侧的无功能韧皮部区域和周皮的结构则类似橡胶树树皮中的砂皮和粗皮。
采割和应用生漆在我国已有数千年的历史(张飞龙,2006; 张飞龙等,2007),经过长时间的生产实践,目前在我国的生漆产区已形成一套成熟的割漆技术,如割漆树龄、割口样式、割口位置的选择以及生漆质量控制方面都有成熟的生产经验。在割漆深度方面,产区实践认为以透过韧皮部割到形成层为宜,漆农谚语“恨皮爱骨”说明了割漆不能伤害“骨头”即木质部(汪明正等,2008)。从本研究中漆树韧皮部的结构来看,漆树韧皮部可分为具功能和无功能韧皮部2部分,其中具功能韧皮部中筛管饱满,乳汁道形态呈多角形或椭圆形,尚未完全发育成熟。因此,在采割生漆时,割口深度应以不伤及具功能韧皮部为宜。如果割得太浅,未达到乳汁道或割断的乳汁道数量较少,生漆流量少,产量低; 如割漆太深,伤及具功能韧皮部筛管和未完全发育成熟乳汁道,甚至伤及形成层和有功能的木质部,不仅影响漆树生长发育,缩短漆树寿命,而且有可能影响下一年的生漆产量。另外,受伤的筛管和导管还会渗出大量水分及其他物质,影响漆液质量。因此可将产区现有的割漆技术加以改良,减小割漆深度,以不割断具功能韧皮部为宜; 采用这种方式割漆,能够有效地保护漆树的正常输导功能和发育状态中的乳汁道,并有利于漆树的生长发育和生漆生产的持续,而且漆树导管、筛管中的水分和其他物质也不会流出混杂到生漆液体中,能保证生漆的质量。
3.2 漆树韧皮部各部分结构的发育规律与生理功能的关系韧皮部的主要生理功能是运输有机养料、合成和贮存次生代谢产物,同时还具有支撑和保护作用。韧皮部的输导功能主要由筛管和伴胞承担,贮存次生代谢产物的功能主要由韧皮薄壁细胞和分泌组织承担,厚壁组织和周皮承担支撑和保护作用。漆树茎干树皮的基本结构类似一般双子叶植物,由维管形成层、次生韧皮部(含具功能和无功能韧皮部)及周皮组成。其中次生韧皮部所占比例最大,次生韧皮部由筛管、伴胞、韧皮薄壁组织、乳汁道、石细胞以及韧皮射线组成。通过韧皮部发育过程的观察表明,各类组成分子分化、发育的时间不同,其差异可能与所承担的生理功能有关。由维管形成层活动产生、分化出的具功能韧皮部中,筛管已发育成熟,占该区域的主要部分,行使输导功能,而韧皮薄壁细胞、乳汁道和韧皮射线尚处于分化过程或幼嫩阶段,未发育成熟。为此,具功能韧皮部应是漆树主要行使养料运输功能的区域。无功能韧皮部中,上述各类组织的细胞都进一步发育变化,并产生新的组织—石细胞。其中,筛管和伴胞发生萎缩,失去输导功能。韧皮薄壁细胞继续发育,其数量和所占比例均逐步增加,最后将其间失去功能的筛管挤毁,在临近周皮处相连成片,其细胞内贮存营养物质。乳汁道发育成熟,分泌和贮存生漆,但在无功能韧皮部外侧,乳汁道的分泌细胞逐渐衰老、萎缩,失去分泌生漆的功能。在具功能韧皮部外侧一定距离的无功能韧皮部中,由一些韧皮薄壁细胞分化出石细胞群,在临近周皮处,石细胞群分布增多,它们增强了树皮的支持和保护作用,以上结构发育特点与多数双子叶木本植物的发育相似,但与葡萄属(Vitis)的茎干树皮不同; 后者当年产生的次生韧皮部的筛管在冬季其筛孔被胼胝质堵塞,失去输导功能,次年春季胼胝质溶解,筛管又恢复输导功能,其筛管可保持生活力多年(Esau,1948),因此,其次生韧皮部难以区分为具功能和无功能2部分。
漆树次生韧皮部中存在大量的石细胞群,目前普遍认为石细胞是由薄壁细胞形成次生壁并沉积大量木质素形成的(Tao et al.,2009),本文研究也表明木质素沉积在漆树石细胞分层的次生壁上。木质素是一类酚类聚合物,主要由香豆酸、松柏醇和芥子醇等单体组成,通常沉积在管状分子和其他植物组织细胞的次生细胞壁上形成厚壁组织以提供机械支持力(Zhong et al.,2000);本文研究发现,具功能韧皮部中无石细胞的产生,石细胞均存在于无功能韧皮部中; 石细胞的产生会增加割漆难度,对生漆生产有一定的负面影响(汪明正等,2008); 漆树韧皮部石细胞群的形成和调控机制有待于进一步研究。
3.3 漆树韧皮部结构发育和季节性变化及其意义树木形成层的活动样式主要有3类: 1)温带和地中海树种,春季恢复活动,夏末秋初进入休眠,形成清晰的年轮; 2)热带荒漠灌木在每年雨季活动,旱季休眠,有或无年轮; 3)生长在有限区域的热带、亚热带树种的形成层全年都处于活动期,有或无年轮(Fahn et al.,1990)。温带地区树木维管形成层活动呈现周期性,春夏季细胞分裂旺盛,秋季减弱,冬季休眠(Derr et al.,1967)。漆树主要分布在温带地区,属于温带经济林木,漆树茎干的维管形成层活动与温带多数双子叶树木相似,呈现上述周期性活动规律。漆树是生漆的原料植物,合成和贮存生漆的结构为乳汁道(胡正海等,1979),漆树韧皮部结构发育研究表明,夏季和初秋季节树皮中成熟乳汁道腔内充满生漆,秋末和冬季减少。因此,在陕西及其周边地区,以6—8月作为采割生漆的季节符合韧皮部及形成层结构的发育规律。
在对植物维管系统的研究发现,当植物导管丧失输导能力以后,导管腔内因水分的丧失而压力下降,在此情况下导管周围的薄壁细胞恢复分裂后会通过纹孔进入导管形成侵填体,从而导致导管堵塞(Sun et al.,2006; 2008),植物筛管中也有类似的情况,筛管中的侵填物质或细胞称为拟侵填体(Lawton,1972),一般情况下侵填体或拟侵填体对导管以及筛管输导功能的破坏作用是不可逆的。漆树乳汁道在韧皮部中是一个多细胞组成的相对封闭的长形腔道,在分泌并贮存生漆的季节,腔道能够保持内外压力的平衡,而在割漆季节结束后,乳汁道内不再分泌并贮存有生漆,此时也会造成腔道内外压力的失衡; 漆树乳汁道在生漆分泌季节结束后产生拟侵填体的结构以保持内外压力的平衡。而在第2年春季,多数乳汁道又重新恢复畅通并贮存有生漆,说明作为拟侵填物的细胞可能已解体。乳汁道拟侵填体是否是通过细胞程序死亡解体,死亡信号是什么以及解体后物质的去向等都是值得深入研究的问题。此外,有关此种季节性变化还有研究表明,在其树皮进入休眠期时,乳汁道腔内产生的拟侵填体细胞不分泌生漆,而产生具有防病、虫作用的醌类物质(张永清,1991),也是一种保护性措施。
| [] | 傅淑颖, 魏朔南, 胡正海. 2007. 漆树各器官中乳汁道的分布与结构特征研究. 西北植物学报, 27(4): 4651–4656. |
| [] | 郝秉中, 吴继林, 云翠英. 1980. 巴西橡胶树有输导功能的韧皮部与采胶的关系. 植物学报, 22(3): 227–231, 310. |
| [] | 胡正海, 王秦生. 1979. 漆树乳汁道的研究. 科学通报(17): 809–812. |
| [] | 汪明正, 刘宏. 2008. 生漆采割技术. 陕西农业科学, 54(2): 223–224. |
| [] | 张飞龙. 2006. 中国漆文化历史渊源研究. 中国生漆, 25(1): 6–20. |
| [] | 张飞龙, 张武桥, 魏朔南. 2007. 中国漆树资源研究及精细化应用. 中国生漆, 26(2): 36–50, 60. |
| [] | 张永清. 1991. 植物次生物质的抗虫作用. 生物学通报(5): 8–9. |
| [] | 张英伯, 郑槐明, 龙瑞芝. 1982. 八种华北树木形成层季节活动及韧皮部与木质部形成的研究. 林业科学, 18(4): 366–453. |
| [] | 张振珏. 1991. 多年生植物韧皮部发育的季节变化和筛分子寿命. 植物学通报, 8(4): 21–25. |
| [] | 张振珏, 陈忠仁, 张永田. 1994a. 黄檀属两种树木形成层的活动周期和次生韧皮部的季节变化. 植物学报, 36(4): 300–304, 332. |
| [] | 张振珏, 林锦仪, 张永田. 1994b. 女贞和白蜡树的树皮结构及次生韧皮部发育的季节变化. 热带亚热带植物学报, 2(3): 28–33. |
| [] | 张振珏, 林锦仪, 陈忠仁. 1997. 四种单宁植物形成层的活动周期和次生韧皮部的季节变化. 云南植物研究, 19(3): 271–274. |
| [] | 赵桂仿, 胡正海. 1990. 漆树乳汁道周围组织的亚显微结构及其与生漆产生关系的研究. 林业科学, 26(1): 17–102. |
| [] | 赵桂仿, 胡正海. 1991. 漆树乳汁道分泌细胞的超微结构及其与生漆产生和分泌关系的研究. 植物学报, 33(1): 26–92. |
| [] | Alfieri F J, Evert R F. 1973. Structure and seasonal development of the secondary phloem in the Pinaceae. Botanical Gazette, 134(1): 17–25. DOI:10.1086/336674 |
| [] | Beaman J H. 1986. Allergenic Asian Anacardiaceae. Clinics in Dermatology, 4(2): 191–203. DOI:10.1016/0738-081X(86)90078-7 |
| [] | Derr W F, Evert R F. 1967. The cambium and seasonal development of the phloem in Robinia pseudoacacia. American Journal of Botany, 54(2): 147–153. DOI:10.2307/2440792 |
| [] | Esau K. 1948. Phloem structure in the grapevine and its seasonal changes. Hilgardia, 18(5): 217–296. DOI:10.3733/hilg.v18n05p217 |
| [] | Ewers F W, Aloni R. 1987. Seasonal secondary growth in needle leaves of Pinus strobus and Pinus brutia. American Journal of Botany, 74(7): 980–987. DOI:10.2307/2443937 |
| [] | Fahn A, Werker E. 1990. Seasonal cambial activity. Taunton, UK:Research Studies Press, 139-157. |
| [] | Hagel J M, Yeung E C, Facchini P J. 2008. Got milk? The secret life of laticifers. Trends in Plant Science, 13(12): 631–639. DOI:10.1016/j.tplants.2008.09.005 |
| [] | Hong D H, Han S B, Lee C W. 1999. Cytotoxicity of urushiols isolated from sap of Korean lacquer tree (Rhus vernicifera Stokes). Archives of Pharmacal Research, 22(6): 638–641. DOI:10.1007/BF02975339 |
| [] | Kim H S, Yeum J H, Choi S W. 2009. Urushiol/polyurethaneurea dispersions and their film properties. Progress in Organic Coatings, 65(3): 341–347. DOI:10.1016/j.porgcoat.2009.02.002 |
| [] | Lawton J R. 1972. Seasonal variations in the secondary phloem of some forest trees from Nigeria Ⅱ. Structure of the phloem. New Phytologist, 71: 335–348. DOI:10.1111/nph.1972.71.issue-2 |
| [] | Samanani N, Alcantara J, Bourgault R. 2006. The role of phloem sieve elements and laticifers in the biosynthesis and accumulation of alkaloids in opium poppy. The Plant Journal, 47(4): 547–563. DOI:10.1111/tpj.2006.47.issue-4 |
| [] | Sun Q, Rost T L, Matthews M A. 2006. Pruning-induced tylose development in stems of current-year shoots of Vitis vinifera(Vitaceae). American Journal of Botany, 93(11): 1567–1576. DOI:10.3732/ajb.93.11.1567 |
| [] | Sun Q, Rost T L, Matthews M A. 2008. Wound-induced vascular occlusions in Vitis vinifera (Vitaceae) : Tyloses in summer and gels in winter. American Journal of Botany, 35(12): 1498–1505. |
| [] | Tao S, Khanizadeh S, Zhang H. 2009. Anatomy, ultrastructure and lignin distribution of stone cells in two Pyrus species. Plant Science, 176(3): 413–419. DOI:10.1016/j.plantsci.2008.12.011 |
| [] | Tucker C M, Evert R F. 1969. Seasonal development of the secondary phloem in Acer negundo. American Journal of Botany, 56(3): 275–284. DOI:10.2307/2440850 |
| [] | Vogl O. 2000. Oriental lacquer, poison ivy, and drying oils. Journal of Polymer Science Part A: Polymer Chemistry, 38(24) : 4327-4335. |
| [] | Webb M. 2000. Lacquer: technology and conservation: a comprehensive guide to the technology and conservation of Asian and European lacquer. Bath, UK: Bath Press, 1-11. |
| [] | Wimmer R, Grabner M. 1997. Effects of climate on vertical resin duct density and radial growth of Norway spruce[Picea abies (L.)Karst.]. Trees-Structure and Function, 11(5): 271–276. |
| [] | Yamauchi Y, Oshima R, Kumanotani J. 1982. Configuration of the olefinic bonds in the heteroolefinicside-chains of japanese lacquer urushiol: separation and identification of components ofdimethylurushiol by means of reductive ozonolysis and highperformance liquid chromatography. Journal of Chromatography A, 243(1): 71–84. DOI:10.1016/S0021-9673(00)88165-0 |
| [] | Zhong R, Ripperger A, Ye Z H. 2000. Ectopic deposition of lignin in the pith of stems of two Arabidopsis mutants. Plant Physiology, 123(1): 59–69. DOI:10.1104/pp.123.1.59 |