 2012, Vol. 48
2012, Vol. 48文章信息
- 刘华波, 王哲, 刘君, 麻丽颖, 王斯琪, 唐岩, 庞晓明
- Liu Huabo, Wang Zhe, Liu Jun, Ma Liying, Wang Siqi, Tang Yan, Pang Xiaoming
- 燕山山脉西伯利亚杏的遗传多样性和遗传结构
- Genetic Diversity and Genetic Structure of Siberian Apricot Populations in the Yan Mountains
- 林业科学, 2012, 48(8): 68-74.
- Scientia Silvae Sinicae, 2012, 48(8): 68-74.
-
文章历史
- 收稿日期:2011-12-06
- 修回日期:2012-02-02
-
作者相关文章



西伯利亚杏(Prunus sibirica),为亚洲特有的生态经济型树种,在我国主要分布在北部和东北部的山区(张加延等,2003)。西伯利亚杏喜光、耐旱、耐寒、耐瘠薄,能够适应各种恶劣的环境条件,是我国丘陵山区阳坡、半阳坡造林绿化的先锋树种(刘桂森等,1990)。西伯利亚杏的种仁不仅是一种传统的干果类食品,而且还是食品、化妆品、制药工业等的重要原料(李洁等,2009),同时,也是开发生物柴油的重要木本油料植物(罗艳等,2007;Gumus et al., 2009;王利兵,2011),因此西伯利亚杏具有非常广阔的开发前景。西伯利亚杏具有自交不亲和性,经过长期的杂交变异和地区间的相互引种,形成了许多变异类型,种质资源极为丰富(李明等,2011a)。目前,人们已经在形态学、生化水平、同工酶等方面对西伯利亚杏特异种质进行了大量的调查研究(张立彬等,1993;2004;赵桂玲,2003;刘明国等,2006;李明等,2011b)。李振江(2007)利用RAPD分子标记对22份来自河北省赤城县的西伯利亚杏特异种质进行了分析;芦宁超(2008)利用RAPD分子标记对吉林省西伯利亚杏的遗传多样性进行了研究。然而上述研究多数是以个体为研究材料,因而不能揭示西伯利亚杏在物种水平上的遗传多样性水平,导致至今西伯利亚杏群体遗传多样性分布资料缺乏。燕山山脉南临华北平原,北接蒙古高原,东濒渤海,西连黄土高原,是华北平原向蒙古高原过渡的区域,有明显的温带大陆性季风气候的特点,冬季寒冷干燥,夏季高温较多雨(娄安如等,2001)。在行政区域上,燕山山脉位于内蒙古东南部、河北北部、辽宁西部三省交汇的区域,是西伯利亚杏分布最为集中的地区(张加延等,2003;王利兵,2010)。此外,燕山山脉丰富的西伯利亚杏资源蕴藏着众多的特异种质,为今后晚花西伯利亚杏选育工作的开展提供了基础(张立彬等,2004)。大量的西伯利亚杏不仅表明其对环境具有较强的适应性,而且在改善燕山山脉地区的生态环境方面具有较大的意义,同时也为这一地区带来可观的经济价值。微卫星分子标记具有重复性好、多态性高和共显性等特性而在植物遗传多样性研究、品种鉴定、分子标记辅助育种等方面得到了广泛的应用(Tautz,1989),因此本研究采用SSR分子标记对燕山山脉地区西伯利亚杏群体的遗传多样性和遗传结构进行了分析,以期为燕山山脉地区西伯利亚杏种质资源的收集和进一步开发利用提供科学依据。
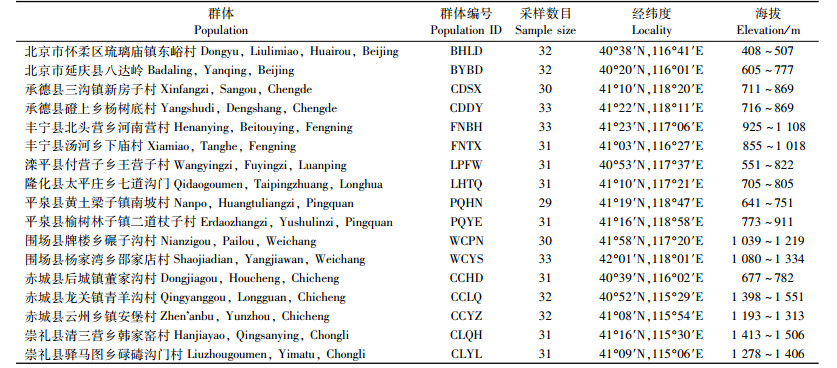
1 材料与方法 1.1 样品采集2011年7,8月份对燕山山脉地区的西伯利亚杏进行了随机采样,在河北北部的承德、张家口及北京的延庆、怀柔采集到17个野生或半野生西伯利亚杏群体,533个个体,具体群体编号、采样地点、样本量等信息见表 1。除LHTQ,PQHN,CCLQ,CCYZ为半野生群体,其余群体均为野生群体特征,采样均尽量覆盖整个群体,每个群体随机选择单株30个左右,相邻株间至少相距50 m左右,并且通过GPS定位系统进行保存。采集的叶片立即放入预装变色硅胶的自封袋内干燥保存,用于DNA的提取。
|
|
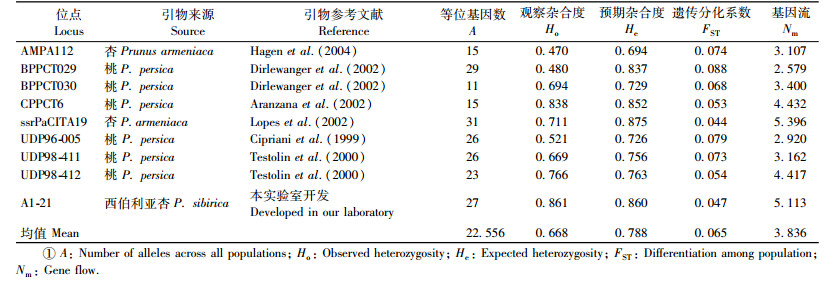
采用改良的CTAB法提取山杏基因组DNA(Doyle et al., 1987)。通过1.0%琼脂糖凝胶电泳和紫外分光光度计检测提取DNA的质量及浓度,DNA样品放于-20 ℃冰箱保存备用。试验选用9对多态性较高的SSR引物(表 2),引物合成时在每一个正引物的5′末端添加一段通用M13序列(5′-TGT AAA ACG ACG GCC AGT-3′)。PCR反应体系:10 ng模板DNA,2×Taq mix 5 μL,正引物(10 μmol·L-1) 0.4 μL,反引物(10 μmol·L-1) 1.6 μL,带有荧光的M13引物(10 μmol·L-1) 1.6 μL,用ddH2O补加至10 μL。参照Schuelke(2000)的扩增程序进行扩增反应,反应程序为:94 ℃ 5 min,[94℃ 30 s,55 ℃ 40 s,72 ℃ 45 s],30个循环,[94 ℃ 30 s,53 ℃ 40 s,72 ℃ 45 s],8个循环,72 ℃ 10 min,10 ℃保存。扩增产物经由DNA分析仪(ABI 3730XL)进行检测,利用Gene-Marker软件(SoftGenetics LLC, USA)读取结果,记录每个位点的片段大小。
|
|
采用GenALEx 6.4(Peakall et al., 2006)软件计算常规的遗传参数:平均每个位点和群体的等位基因数(分别为A,Na)、平均有效等位基因数(Ne)、观察杂合度(Ho)、预期杂合度(He)、Shannon多样性指数(I)、近交系数(F)。应用FSTAT 2.9.3软件(Goudet, 2001)计算两两群体间的遗传分化系数FST。根据公式Nm≈(1-FST)/4FST(Slatkin et al., 1989)估计群体间基因流。由Arlequin 3.5软件(Excoffier et al., 2010)对西伯利亚杏群体遗传变异进行分子方差分析(AMOVA),估测遗传变异在群体内和群体间的分配。由FST/(1-FST)(Rousset, 1997)估计群体间的遗传距离,由经纬度估计群体间的Euclidean地理距离(km),采用IBD3.21(http://ibdws.sdsu.edu/, Jensen et al., 2005)进行10 000个随机矩阵的Mantel检测,分析地理距离和遗传距离的相关性。利用GenALEx 6.4计算Nei's无偏遗传一致度(I)和遗传距离(D),运用NTSYS-pc 2.10e软件(Rohlf, 1994)聚类分析。
2 结果与分析 2.1 群体遗传多样性9对微卫星引物在燕山山脉17个西伯利亚杏群体中共检测到203个等位基因,各个位点的等位基因数为11个(BPPCT030)到31个(ssrPaCITA19),平均每个位点的等位基因数(A)为22.556个(表 2)。所有位点在17个西伯利亚杏群体中均表现多态性(表 3),多态性位点百分率为100%。各群体每位点的平均等位基因数(Na)为8.667~13.556,最高的群体为北京八达岭(BYBD),最低的群体为崇礼清三营(CLQH)和崇礼驿马图(CLYL)。各群体平均有效等位基因数(Ne)为4.188(CLYL)~7.255(PQYE)。各群体的平均观察杂合度(Ho)为0.587(CLQH)~0.761(WCPN),平均预期杂合度(He)为0.727(CLYL)~0.837(BYBD)。Shannon多样性指数(I)为1.633(CLYL)~2.159(BYBD)。对野生群体与半野生群体的遗传多样性参数的均值进行了比较,有效等位基因数(Ne)、预期杂合度(He)和Shannon信息指数(I)分别为5.824,0.792,1.906;5.354,0.775,1.826。半野生群体PQHN的遗传多样性水平(Ne=6.497;He=0.813;I =2.006)高于野生群体遗传多样性的均值,表现出高水平的遗传多样性;其余3个半野生群体的遗传多样性水平低于野生群体遗传多样性的均值,但高于野生群体CLYL。近交系数(F)为0.061(WCPN)~0.221(CLQH和FNTX),均值为0.155。根据平均有效等位基因数(Ne)、平均预期杂合度(He)、Shannon多样性指数(I),遗传多样性水平最高的群体为北京八达岭(BYBD)、平泉榆树林子(PQYE),最低的群体为崇礼驿马图(CLYL)和崇礼清三营(CLQH)。
|
|

燕山山脉17个西伯利亚杏群体总的遗传分化系数FST为0.065,遗传分化最低的2个群体为PQYE和PQHN(0.005 9),最高的2个群体为PQYE和CLYL(0.099 4);群体间总的基因流为3.836,PQYE和CLYL间基因流最小(2.265 1),PQYE和PQHN间基因流最大(42.122 9)。AMOVA分析表明,燕山山脉西伯利亚杏遗传变异主要存在于群体内个体间(95.62%),群体间的遗传变异只有4.38%,群体内和群体间的变异均极显著(表 4)。
|
|
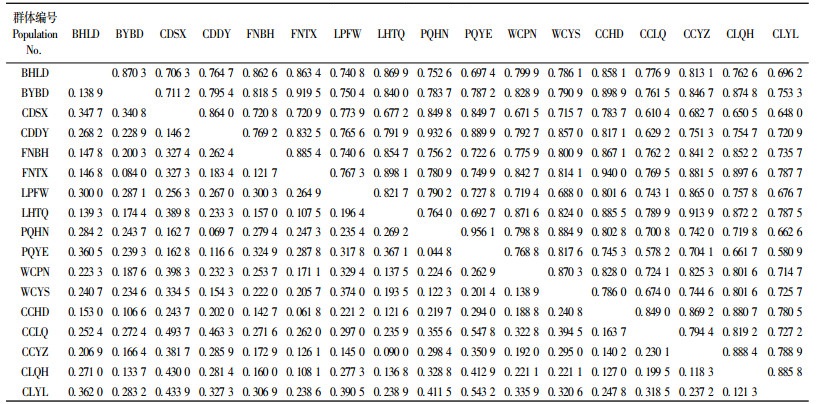
Mantel检测结果表明,遗传距离与地理距离之间存在显著相关(r=0.589 4,P < 0.000 1)(图 1)。根据Nei's方法计算出西伯利亚杏两两群体间的Nei's无偏遗传距离(D)和遗传一致度(I)见表 5。PQYE和CCLQ群体间的Nei's无偏遗传距离(D)最大(0.547 8),遗传一致度最低(0.578 2)。PQYE和PQHN群体间的Nei's无偏遗传距离(D)最小(0.044 8),遗传一致度最高(0.956 1)。UPGMA聚类结果(图 2)显示:平泉县内的2个群体和承德县内的2个群体聚在一起,最后同滦平县内的群体聚在一起,构成了第1个类群;赤城龙关群体单独与其余所有群体聚在一起构成了第2个类群。大部分地理位置较近的群体都聚在了一起,这也进一步验证了Mantel检测的结果。

|
图 1 遗传距离与地理距离的相关性 Fig.1 Correlation between genetic distance and geographical distance of all sampled populations of Siberian apricot |
|
|

|
图 2 西伯利亚杏群体Nei's无偏遗传距离UPGMA聚类图 Fig.2 UPGMA dendrogram of Siberian apricot populations based on Nei's unbiased genetic distance |
芦宁超(2008)用RAPD分子标记对吉林省西伯利亚杏的遗传多样性进行研究,16个引物共扩增出124条带,多态性带百分率P为90.32%,本研究应用SSR分子标记对燕山山脉西伯利亚杏群体进行遗传多样性及遗传结构的分析,多态性带百分率P为100%,表明燕山山脉西伯利亚杏群体具有非常丰富的遗传多样性。何天明(2005)利用SSR分子标记分别对伊犁河谷地区野杏(Prunus armeniaca)和普通杏品种的遗传多样性水平进行研究,多态性带百分率P、期望杂合度He和Shannon信息指数I分别为82.4%,0.287和0.428(野杏);85.3%,0.264和0.408(普通杏),明显低于燕山山脉西伯利亚杏的遗传多样性水平(P=100%,He=0.788,I=1.887)(表 3),也表明燕山山脉西伯利亚杏群体具有较高的遗传多样性水平。与其他多年生木本植物天然群体的遗传多样性相比(郑健等,2008;谢一青等,2008;田华等,2009;左丹丹等,2009),燕山山脉地区西伯利亚杏群体仍然表现为较高的遗传多样性水平。
多数学者普遍认为分布广、多年生、异交的物种遗传多样性水平较高(Hamrick,1990;郑健等,2008)。西伯利亚杏广泛分布在我国三北地区的多个省市,对各种恶劣的环境具有较高的适应性。刘桂森等(1990)研究表明西伯利亚杏自花结实的能力比较低,表现出自交不亲和性,株间自然授粉是其结实的主要途径。刘黎明(2010)对新疆野杏生殖生态的研究表明,野杏的主要访花昆虫为蜂类、蝇类和极少数的蝶类,虽然目前还缺乏对西伯利亚杏的传粉昆虫的直接调查研究,但同新疆野杏应具有相似的传粉者。本文的研究结果表明西伯利亚杏群体具有较高杂合度(Ho=0.668, He=0.788)和较低的近交系数F(0.155),因此推断西伯利亚杏的交配系统主要为异交。异交和虫媒花这样的繁育特性对于保持较高的遗传多样性是有利的(汪书丽等,2007)。此外,西伯利亚杏主要是通过核果进行繁殖,果实成熟后自然开裂脱落。马庆亮(2010)的研究中发现,大林姬鼠(Apodemus peninsulae)、社鼠(Niviventer confucianus)、黑线姬鼠(Apodemus agrarius)等啮齿动物会对西伯利亚杏进行取食,同栓皮栎(Quercus variabilis)种子相比,西伯利亚杏种子的扩散速率更大,搬运距离更远,且啮齿动物更倾向于贮藏西伯利亚杏种子。啮齿动物这种特殊的贮食策略给西伯利亚杏提供了远距离基因交流的可能性,也带来了增加遗传多样性的原因。另外,多态位点比率在引物之间存在着差异,本研究所用的标记为从多个标记中筛选得到的多态性标记。以上因素都可能是造成西伯利亚杏群体遗传多样性较高的原因。通过对西伯利亚杏野生群体与半野生群体的遗传多样性参数的比较,本研究发现半野生群体的遗传多样性水平与野生群体的差异较小,半野生群体同样可以保持较高的遗传多样性水平,不同的半野生群体间的遗传多样性差异明显,这可能是与选取的种源群体的遗传多样性水平有关。
3.2 群体遗传分化根据Wright(1978)的建议,FST值在0~0.05时,群体间的遗传分化很小;FST为0.05~0.15,群体间存在中等程度的遗传分化;FST为0.15~0.25,群体间遗传分化较大;FST大于0.25,群体间有很大的遗传分化。燕山山脉地区西伯利亚杏群体间的遗传分化系数FST为0.065,表明其群体间存在中等程度的遗传分化,并且93.5%的遗传变异存在于群体内。这与分子方差分析(AMOVA)得出的大部分遗传变异存在于群体内(95.62%),只有一小部分变异存在于群体间(4.38%)(表 4)的结果相吻合,与伊犁野杏86.3%(He et al., 2007)、普通杏85.3%(苑兆和,2007),以及其他多年生木本植物的变异规律相符(刘军等,2008;乔小燕等,2011)。
Hamrick(1990)统计了165属449种裸子植物和被子植物的研究结果,认为基因交流程度和繁育系统方式是影响群体遗传结构的最主要因素。Wright(1978)认为适中的基因流可以防止群体遗传分化的发生。根据FST估算西伯利亚杏群体间的基因流为3.836,与花楸树(Sorbus pohuashanensis)上检测到的基因流(3.047 2)比较接近(郑健等,2008),较高水平的基因流表明遗传漂变不是西伯利亚杏群体遗传分化的主要影响因素,这可能与其繁育系统主要为异交,传粉借助昆虫以及种子可被啮齿动物的取食而扩散有关。此外,西伯利亚杏作为我国北方生态林建设重要的树种,地域间的相互引种以及野生群体与半野生群体的混杂生长都有可能是增加群体间基因流的原因。Mantel检测结果也显示群体遗传距离与地理距离间存在显著相关性(r=0.589 4,P < 0.000 1),进一步表明了燕山地区西伯利亚杏群体间的基因流是影响群体遗传结构的重要因素。
3.3 西伯利亚杏特异种质资源的收集和保护策略本研究揭示西伯利亚杏具有较高的遗传多样性,在野外调查中发现燕山地区西伯利亚杏存在着丰富的遗传变异,特异种质资源丰富。平泉榆树林子镇发现多株绿萼西伯利亚杏以及大花、粉花类型,北京怀柔地区发现双柱头比例较高的类型,以及延庆地区的雌蕊退化类型。丰富的特异种质资源可为西伯利亚杏生态林的营建提供丰富的变异来源,因此,对于分布在各个群体的特异种质要加大力度进行异地保存和利用。基于燕山山脉西伯利亚杏群体间遗传变异系数较低,遗传变异主要存在于群体内这一研究结果,对少数地区的群体进行采集即可代表大部分地区的遗传变异,但每个群体应采集尽可能多的植株,以涵盖该物种的基因库,同时也要注意其他地区的特异个体。北京八达岭群体的遗传多样性最高,但以往较少涉及,今后应给以更多的关注。
本研究采用微卫星分子标记技术对燕山山脉地区的西伯利亚杏群体进行了遗传多样性和遗传结构的研究,可以为西伯利亚杏资源的合理开发与保护提供科学依据。燕山山脉地区是西伯利亚杏分布集中的地区,因而应该得到优先保护。另外,为全面揭示西伯利亚杏群体的多样性水平和群体结构,还需要从更多的地区进行样品采集和分析。
| [] | 何天明. 2005. 中国普通杏(Prunus armeniaca)种质资源遗传多样性及紫杏(P. dasycarpa)起源研究. 山东农业大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10434-2006100760.htm |
| [] | 李洁, 李旺, 彭艳芳, 等. 2009. 野生山杏资源的综合开发和利用. 河北林业科技(3): 38–40. |
| [] | 李明, 赵忠, 杨吉安, 等. 2011a. 黄土高原山杏种质资源分类研究. 西北林学院学报, 26(1): 8–12. |
| [] | 李明, 赵忠, 杨吉安, 等. 2011b. 黄土高原不同县域山杏种质资源遗传多样性研究. 西北农林大学学报, 39(2): 143–149. |
| [] | 李振江. 2007. 山杏良种资源的RAPD分析. 河北农业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-11920-2007163492.htm |
| [] | 刘军, 陈益泰, 孙宗修, 等. 2008. 基于空间自相关分析研究毛红椿天然居群的空间遗传结构. 林业科学, 44(6): 45–52. DOI:10.11707/j.1001-7488.20080608 |
| [] | 刘黎明. 2010. 新疆野杏生殖生态学初步研究. 新疆农业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10758-1011040971.htm |
| [] | 刘桂森, 张立彬. 1990. 野生山杏的结实特征和果实发育动态的研究. 河北农业技术师范学院学报, 4(4): 10–16. |
| [] | 刘明国, 赵桂玲, 董胜君. 2006. 山杏种内POD同工酶及种子可溶性蛋白分析. 沈阳农业大学学报, 37(4): 582–586. |
| [] | 娄安如, 刘文华. 2001. 燕山山脉植物群落的间接梯度分析与数量分类. 北京师范大学学报, 37(3): 391–395. |
| [] | 芦宁超. 2008. 吉林省野生西伯利亚杏种质资源遗传多样性研究. 吉林农业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10193-2009162692.htm |
| [] | 罗艳, 刘梅. 2007. 开发木本油料植物作为生物柴油原料的研究. 中国生物工程杂志, 27(7): 68–74. |
| [] | 马庆亮. 2010. 王屋山地区啮齿动物对林木种子的扩散与贮藏. 郑州大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10459-1011017363.htm |
| [] | 乔小燕, 乔婷婷, 周炎花, 等. 2011. 基于EST-SSR的广东与广西茶树资源遗传结构和遗传分化比较分析. 中国农业科学, 44(16): 3297–3311. DOI:10.3864/j.issn.0578-1752.2011.16.002 |
| [] | 田华, 康明, 李丽, 等. 2009. 中国板栗自然居群微卫星(SSR)遗传多样性. 生物多样性, 17(3): 296–302. |
| [] | 王利兵. 2010. 我国3种杏的地理分布及其植物学性状. 林业科学研究, 23(3): 435–439. |
| [] | 王利兵. 2011. 三种山杏种子生物柴油特征评价. 农业工程学报, 27(S1): 138–142. |
| [] | 汪书丽, 马巧明. 2007. 中国木棉居群的遗传多样性. 云南植物研究, 29(5): 529–536. |
| [] | 谢一青, 李志真, 黄儒珠, 等. 2008. 武夷山不同海拔光皮桦种群遗传多样性及其与生态因子的相关性. 林业科学, 44(3): 50–55. DOI:10.11707/j.1001-7488.20080313 |
| [] | 苑兆和. 2007. 杏属植物种质资源分子系统学研究. 山东农业大学博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10434-2007135653.htm |
| [] | 张加延, 张钊. 2003. 中国果树志:杏卷. 北京:中国林业出版社: 32–33. |
| [] | 张立彬, 刘桂森. 1993. 野生山杏性状变异性及其应用的研究. 河北农业技术师范学院学报, 7(3): 1–6. |
| [] | 张立彬, 王同坤, 刘桂森, 等. 2008. 燕山野生山杏种内变异//中国园艺学会. 第十一次全国李杏会议论文集. 成都: 157-159. |
| [] | 赵桂玲. 2003. 半干旱地区山杏种质资源调查及遗传特质调查. 沈阳农业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10157-2003120989.htm |
| [] | 郑健, 郑勇奇, 张川红, 等. 2008. 花楸树天然群体的遗传多样性研究. 生物多样性, 16(6): 562–569. |
| [] | 左丹丹, 赵海涛, 刘春, 等. 2009. 蜡梅天然群体遗传多样性的SRAP标记分析. 园艺学报, 36(8): 1197–1202. |
| [] | Aranzana M J, Garcia-Mas J, Carbó J, et al. 2002. Development and variability analysis of microsatellite markers in peach. Plant Breeding, 121(1): 87–92. DOI:10.1046/j.1439-0523.2002.00656.x |
| [] | Cipriani G, Lot G, Huang W G, et al. 1999. AC/GT and AG/CT microsatellite repeats in peach(Prunus persica(L.) Batsch): Isolation, characterization and cross-species amplification in Prunus. Theor Appl Genet, 99(1/2): 65–72. |
| [] | Dirlewanger E, Cosson P, Tavaud M, et al. 2002. Development of microsatellite markers in peach(Prunus persica(L.) Batsch) and their use in genetic diversity analysis in peach and sweet cherry(Prunus avium L.). Theor Appl Genet, 105(1): 127–138. DOI:10.1007/s00122-002-0867-7 |
| [] | Doyle J J, Doyle J L. 1987. A rapid DNA isolation procedure for small quantities of leaf tissue. Phytochem Bull, 19: 11–15. |
| [] | Excoffier L, Lischer H E L. 2010. Arlequin suite ver3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources, 10(3): 564–567. DOI:10.1111/men.2010.10.issue-3 |
| [] | Goudet J. 2001. FSTAT, A Program to Estimate and Test Gene Diversities and Fixation Indices(version 2.9.3). http://www.unil.ch/izea/softwares/fstat.html. |
| [] | Gumus M, Kasifoglu S. 2009. Performance and emission evaluation of a compression ignition engine using a biodiesel(apricot seed kernel oil methyl ester) and its blends with diesel fuel. Biom Bioe, 34(1): 134–139. |
| [] | Hagen L S, Chaib J, Fady B, et al. 2004. Genomic and cDNA microsatellites from apricot(Prunus armeniaca L.). Molecular Ecology Notes, 4(4): 742–745. DOI:10.1111/men.2004.4.issue-4 |
| [] | Hamrick J K. 1990. Isozymes and the analysis of genetic structure in plant populations// Soltis D E, Soltis P S. Isozymes in Plant Biology. London: Chapman and Hall, 87-105. |
| [] | He T M, Chen X S, Xu Z, et al. 2007. Using SSR markers to determine the population genetic structure of wild apricot(Prunus armeniaca L.) in the Ily Valley of West China. Genetic Resources and Crop Evolution, 54(3): 563–572. DOI:10.1007/s10722-006-0013-5 |
| [] | Jensen J L, Bohonak A J, Kelley S T. 2005. Isolation by distance, web service. BMC Genetics, 6: 13. |
| [] | Lopes M S, Sefc K M, Laimer M, et al. 2002. Identification of microsatellite loci in apricot. Molecular Ecology Notes, 2(1): 24–26. DOI:10.1046/j.1471-8286.2002.00132.x |
| [] | Peakall R, Smouse P E. 2006. GENALEX 6: genetic analysis in Excel.Population genetic software for teaching and research. Molecular Ecology Notes, 6(1): 288–295. DOI:10.1111/men.2006.6.issue-1 |
| [] | Rohlf F J. 1994.NTSYS-PC: Numerical Taxonomy and Multivariate Analysis System Version 1.80. Setauket New York: Distribution by Exeter SoftWare. http://www.scienceopen.com/review?vid=1866d74a-f3c6-4fdc-84ac-495ddd8d4fcf |
| [] | Rousset F. 1997. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genetics, 145(4): 1219–1228. |
| [] | Schuelke M. 2000. An economic method for the fluorescent labeling of PCR fragments. Nat Biotechnol, 18(2): 233–234. DOI:10.1038/72708 |
| [] | Slatkin M, Barton N H. 1989. A comparison of three indirect methods for estimating average levels of gene flow. Evolution, 43(7): 1349–1368. DOI:10.1111/evo.1989.43.issue-7 |
| [] | Tautz D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucl Acids Res, 17(16): 6463–6471. DOI:10.1093/nar/17.16.6463 |
| [] | Testolin R, Marrazzo T, Cipriani G, et al. 2000. Microsatellite DNA in peach(Prunus persica L.Batsch) and its use in fingerprinting and testing the genetic origin of cultivars. Genome, 43(3): 512–520. DOI:10.1139/g00-010 |
| [] | Wright S. 1978. Evolution and the genetics of populations, Vol.4. Variability within and among natural populations. Chicago: University of Vhicago Press. http://www.jstor.org/stable/2529965 |