 2012, Vol. 48
2012, Vol. 48文章信息
- 汤孟平, 娄明华, 陈永刚, 徐文兵, 赵明水
- Tang Mengping, Lou Minghua, Chen Yonggang, Xu Wenbing, Zhao Mingshui
- 不同混交度指数的比较分析
- Comparative Analyses on Different Mingling Indices
- 林业科学, 2012, 48(8): 46-53.
- Scientia Silvae Sinicae, 2012, 48(8): 46-53.
-
文章历史
- 收稿日期:2011-09-18
- 修回日期:2012-03-13
-
作者相关文章


2. 浙江省森林生态系统碳循环与固碳减排重点实验室 临安 311300;
3. 天目山国家级自然保护区管理局 临安 311300
2. Zhejiang Provincial Key Laboratory of Carbon Cycling in Forest Ecosystems and Carbon Sequestration Lin'an 311300;
3. Management office, National Nature Reserve of Tianmu Mountain Lin'an 311300
生物多样性是国际社会关注的热点问题(马克平,1993),生物多样性保护已成为森林可持续经营的一个重要目标(雷相东等,2002)。生物多样的测度包括从基因到生态系统多个等级。在林分尺度,常用树种多样性表示生物多样性(Lexerød et al., 2006),林分内树种越多,生物多样性就越高,林分就越稳定。
用树种多样性表示林分生物多样性尚有不足之处。树种多样性仅反映树种丰富度,并没考虑树种之间空间隔离关系的多样性,它是对生物多样性的非空间表述。事实上,为定量描述树种之间的空间隔离关系,已提出了多种方法。Peilou(1961)提出分隔指数,可以分析2个种的分隔程度,但对多个种也只能进行两两比较(张金屯,1995)。Gadow等(1992)提出了多个树种空间隔离关系的混交度,对描述树种空间分布多样性具有历史意义。为便于与其他混交度比较,不妨把Gadow等(1992)提出的混交度称之为简单混交度,即在空间结构单元中,对象木的最近邻木与对象木不属同种的个体所占的比例。空间结构单元是由林分中任意一株树木即对象木与其周围最近邻木所构成。
由于Gadow等(1992)提出的混交度仅考虑对象木与最近邻木之间的树种隔离关系,并没有考虑最近邻木之间的树种异同,因此对空间结构单元树种隔离程度的反映是不完整的。汤孟平等(2004)提出了树种多样性混交度。此后,惠刚盈等(2008)指出了树种多样性混交度的不足,认为树种多样性混交度混淆了4个邻体中有3株树种相同(图 1A)和4个邻体中有2株树种相同(图 1B)的混交度。图 1中不同符号表示不同树种,圆表示对象木的树种,三角形和正方形表示最近邻木的树种。为此,惠刚盈等(2008)提出了物种空间状态的概念。

|
图 1 2种不同的混交结构单元 Fig.1 Two different mixed structure units |
事实上,物种空间状态也没有解决树种多样性混交度存在的问题。对图 1中2种不同的混交结构单元计算混交度可知,树种多样性混交度均为0.5,物种空间状态均为0.6,后者大于前者,说明物种空间状态对树种空间隔离程度的灵敏性有所提高。但是,分别采用树种多样性混交度或者物种空间状态,计算得出2种不同混交结构单元的混交度都是相同的,说明树种多样性混交度和物种空间状态都不能区分这2种不同的混交结构。原因在于用最近邻木(树种多样性混交度)或空间结构单元(物种空间状态)的树种数比例来描述最近邻木之间的树种隔离程度是不准确的,因为相同的树种数比例,可能存在不同的树种隔离关系。如图 1A和图 1B所示,虽然2种混交结构单元的树种数比例相同,具体讲当考虑最近邻木时,均为2/4,当考虑空间结构单元时,均为3/5,但它们的树种隔离关系并不相同,而计算混交度结果却相同,这说明没有把不同的树种隔离关系区分开来,因此,不能用树种数比例代替最近邻木相互之间的树种隔离关系。树种数比例可以表达树种多样性,但不能同时表达树种隔离关系。
根据现有混交度的研究不难发现,混交度本质上是定量描述林分空间结构单元中树种隔离程度的指数,它取决于树种多样性和树种空间隔离关系。树种多样性是树种空间隔离的基础,树种越多,不同树种之间相互隔离的可能性越大。树种空间隔离关系包括对象木与最近邻木之间的树种隔离和最近邻木相互之间的树种隔离。但是,现有混交度存在的共同问题是对空间结构单元的树种隔离关系表达不完整,导致不同混交结构具有相同混交度结果。鉴于此,本文提出了一个新的混交度即全混交度。全混交度全面考虑空间结构单元的树种隔离关系和树种多样性,可以避免出现不同混交结构具有相同混交度的现象。本研究以天目山常绿阔叶林数据为例,应用基于GIS的Voronoi图分析功能对不同混交度指数进行比较分析。
1 材料与方法 1.1 全混交度全混交度全面考虑对象木与最近邻木之间以及最近邻木相互之间的树种隔离关系,同时兼顾树种多样性。树种多样性不仅考虑树种数,还考虑不同树种所占比例的均匀度。为此,引入生物多样性Simpson指数(Simpson, 1949; Shimatani, 2001; Onaindiaa et al., 2004)来描述树种多样性,以提高树种多样性的区分度。全混交度的计算公式为:
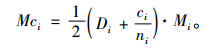
|
(1) |
式中:Mci为第i空间结构单元中对象木的全混交度;Mi是简单混交度,


为了比较全混交度与简单混交度、树种多样性混交度和物种空间状态的差异,在图 1中2种不同混交结构单元基础上,再假定3种典型的混交结构单元(图 2)。图 2A是为了与图 1B进行比较,二者均有3个树种,且各树种株数也相同,唯一区别是4个最近邻木的相互隔离关系不同。图 2B和图 2C则代表树种各不相同的2种混交结构单元,图 2B有5个不同树种,图 2C有6个不同树种,这是假定最近邻木株数可变(现实中常如此)前提下的2种极端树种隔离情形。上述5个混交结构单元的不同混交度计算结果见表 1。

|
图 2 3种典型的混交结构单元 Fig.2 Three typical mixed structure units |
|
|
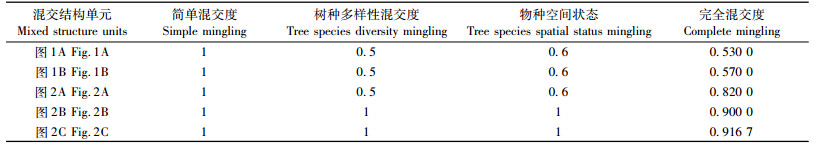
由表 1可见,简单混交度根本不能区分图 1和图 2共5种不同混交结构单元的树种隔离程度。另外,树种多样性混交度和物种空间状态也只有较弱的分辨力,甚至不能区分图 1B和图 2A 2个树种组成相同、但最近邻木相互隔离关系不同的空间结构单元。全混交度则具有最强的树种隔离程度分辨能力,完全可以区分这5种不同的混交结构单元。因此,全混交度对空间结构单元的树种多样性和树种空间隔离关系具有最准确的表达,是对林分生物多样性和林分结构复杂性的一种有效空间表述。
1.2 各混交度之间的数学关系下面将证明各混交度之间存在的数学关系。Mi,Mti,Msi分别为第i空间结构单元的简单混交度、树种多样性混交度和物种空间状态,计算公式见有关文献(Gadow et al., 1992;汤孟平等,2004;惠刚盈等,2008;Hui et al., 2011)。Mi,Mti,Msi和Mci的取值范围均为[0,1],以下不再说明。设ti为第i空间结构单元对象木的相邻木中不同树种数,其他符号同式(1)。
1.2.1 简单混交度与树种多样性混交度、物种空间状态或全混交度的关系定理1 简单混交度大于或等于树种多样性混交度、物种空间状态和全混交度,即Mi≥Mti,Mi≥Msi,Mi≥Mci。
证明 先证明Mi≥Mti。根据简单混交度和树种多样性混交度计算公式,有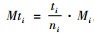
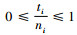
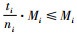
再证明Mi≥Mci。因为0≤Di≤1和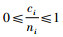
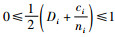

定理2 当空间结构单元的树种数等于最近邻木的树种数时,树种多样性混交度大于或等于物种空间状态,即当ti=si时,Mti≥Msi,当且仅当ti=si=1时,Mti=Msi=0;当空间结构单元的树种数比最近邻木的树种数多1个,或者对象木的树种与最近邻木的树种均不相同时,树种多样性混交度小于物种空间状态,即当ti+1=si时,Mti≤Msi,当且仅当ti=ni时,Mti=Msi=1。
证明 先证第一个结论。当ti=si时,因为ti,si和ni都是正数,所以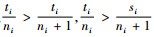

再证第二个结论。当ti+1=si时,则ti < si。因为ti,si和ni都是正数,且ti≤ni,根据不等式的性质,有
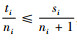
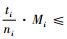
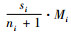
全混交度明显不同于树种多样性混交度和物种空间状态,它们之间的关系十分复杂。根据ci,ti和si之间的关系以及树种多样性,可分为5种关系,分别见定理3,4,5,6和7。
定理3 当空间结构单元的树种数和最近邻木的树种数均为1时,即ti=si=1,则Mti=Msi=Mci=0。
证明 已知ti=si=1,则Mi=0,根据树种多样性混交度、物种空间状态的计算公式和式(1),有Mti=Msi=Mci=0。
定理4 当空间结构单元的树种数和最近邻木的树种数相等,也等于最近邻木中成对相邻木非同种的个数,但不等于1时,即ti=si=ci≠1:
1) 如果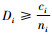
2) 如果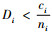
证明 情形1):已知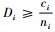
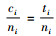
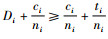
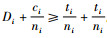

情形2):已知

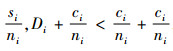
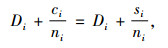
|
(2) |

|
(3) |
当空间结构单元的株数按树种分布最不均匀时,即某一个树种的株数最多、其余树种的株数均为1时,Di达到最小值Dmin:

|
(4) |
根据式(4),必有Di≥Dmin。进一步有
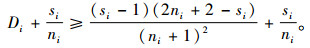
|
(5) |
把式(5)两边减去同一项
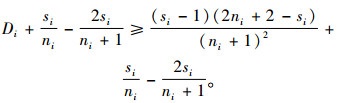
|
(6) |
已知ci=ti=si≠1,则si≥2,且ni+1-si≥0。则式(6)的右端项
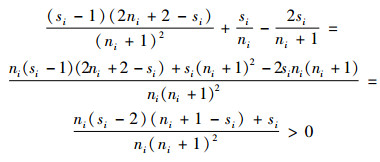
|
因此,根据式(6),有

|
(7) |
把式(3)与(7)相结合,有

|
(8) |
根据式(8),有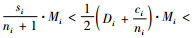

定理5 当空间结构单元的树种数和最近邻木的树种数相等,且成对相邻木非同种的个数至少比最近邻木的树种数多1,即ti=si≠1,ci≥ti+1时,Mci > Mti > Msi。
证明 已知ci≥ti+1,则ci-ti≥1。已知ti=si≠1,且ni≥ci。所以,ni-ti≥1,ni-si≥1。根据式(4),有

|
(9) |
因为式(9)右端项
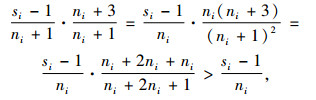
|
所以
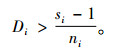
|
(10) |
把式(10)两边加上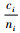

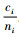
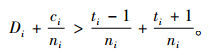
|
(11) |
把式(11)两边除以2,得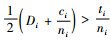
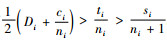
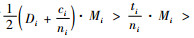

定理6 当空间结构单元的树种数比最近邻木的树种数多1个,且最近邻木中成对相邻木非同种的个数等于最近邻木的树种数,即si=ti+1,ci=ti时:
1) 如果
2) 如果
证明 情形1):已知si=ti+1,ci=ti,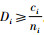



情形2):已知si=ti+1,ci=ti。当
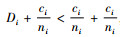

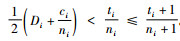
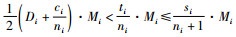
定理7 当空间结构单元的树种数比最近邻木的树种数多1个,且最近邻木中成对相邻木非同种的个数至少比最近邻木的树种数多1个,即si=ti+1,ci≥ti+1时,Mci>Msi≥Mti。
证明 已知si=ti+1,ci≥ti+1。所以,ci≥si,
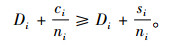
|
(12) |
根据式(4),则式(12)的右端项
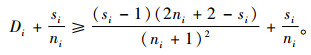
|
(13) |
因为ti≥1,si=ti+1,则si≥2,且ni+1-si≥0。所以式(7)成立,并且有

|
(14) |
根据式(14),有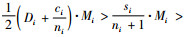
浙江天目山国家级自然保护区位于浙江西北部临安市境内的西天目山(119°23′47″—119°28′27″E, 30°18′30″—30°24′55″N),总面积1 050 hm2。年均气温8.8~14.8 ℃;≥10 ℃积温2 500~5 100 ℃;年降水量1 390~1 870 mm,相对湿度76%~81%。保护区地处中亚热带北缘向北亚热带过渡的地带,受海洋暖湿气候影响,温暖湿润,雨量充沛,森林植被十分茂盛。植被分布有明显的垂直界限,自山麓到山顶垂直带谱为:海拔870 m以下为常绿阔叶林;870~1 100 m为常绿、落叶阔叶混交林;1 100~1 380 m为落叶阔叶林;1 380~1 500 m为落叶矮林(汤孟平等,2007)。
2005年,在保护区内选择典型的常绿阔叶林,设置大小为100 m×100 m的1个大型固定标准地,标准地中心海拔630 m,主坡向南坡。用相邻格子调查方法,把标准地划分为100个10 m×10 m的调查单元,对每个调查单元进行每木调查,并采用激光对中全站仪(徕卡TCR702Xrange)测定每株树木基部三维坐标。2010年,对该标准地进行复查,在每个调查单元内,对胸径≥5 cm的树木进行每木调查,测定每株树木的胸径、树高、枝下高和冠幅等因子。对达到起测径阶的树木,还要记录树种,测定树木基部三维坐标。本研究用2010年复查数据作为基础数据。
1.4 空间结构单元与边缘矫正空间结构单元是森林空间结构分析的基本单位,由对象木和最近邻木组成。对象木是标准地内任意一株树木,最近邻木采用基于GIS的Voronoi图分析方法确定(汤孟平等,2009)。为消除标准地边缘的影响,必须进行边缘矫正。采用缓冲区方法进行边缘矫正,即由标准地的每条边向固定标准地内部水平距离5 m的范围作为缓冲区。在标准地中,除缓冲区外的其余部分称为矫正标准地,矫正标准地大小为90 m×90 m。当计算混交度时,仅把矫正标准地内的全部树木作为对象木。
2 结果与讨论基于GIS进行程序设计,计算每株对象木的4种混交度以及林分平均混交度,并绘制混交度之间的关系图(图 3、图 4)。可以看出,各混交度之间有以下几个明显关系和特点。

|
图 3 不同混交度的关系 Fig.3 Relationship between different mingling |

|
图 4 林分平均混交度 Fig.4 Mean mingling of the stand |
1) 图 3A、图 3B和图 3C反映简单混交度与其他混交度之间的关系(定理1)。从坐标系原点出发的第一象限角平分线表示混交度相等的直线(下同)。可见,所有点分布在角平分线及以下区域,表明简单混交度大于或等于树种多样性混交度(图 3A)、物种空间状态(图 3B)和全混交度(图 3C)。林分平均混交度也显示了同样的大小关系特征(图 4),这验证了各混交度之间关系定理1。说明,简单混交度没有考虑最近邻木相互之间的树种隔离关系以及树种多样性,从而过高估计树种隔离程度。
2) 图 3D反映树种多样性混交度与物种空间状态之间的关系(定理2)。可以看出,离散点被分为上、下2部分,并呈现2个分离的线性关系趋势。下面部分表示当ti=si时,树种多样性混交度大于或等于物种空间状态,仅当ti=si=1时,2种混交度相等且为0,这验证了关系定理2的第1个结论。实际上,当ti=si=1时,全混交度也等于0,这验证了定理3。上面部分表示当ti+1=si时,树种多样性混交度小于或等于物种空间状态,仅当ti=ni时,2种混交度相等且为1,这验证了关系定理2的第2个结论。由于树种多样性混交度与物种空间状态之间存在分离的线性关系,说明对给定的空间结构单元,二者的大小不仅与树种隔离有关,还受空间结构单元的树种数和最近邻木的树种数是否相等的影响,结果出现波动,不便于独立准确描述空间结构单元的树种隔离关系。从平均混交度来看,平均树种多样性混交度 > 平均物种空间状态混交度(图 4),但这不是一般性结论。
3) 图 3E~图 3J反映全混交度与树种多样性混交度和物种空间状态之间的关系(定理4~定理7)。图 3E和图 3F分别验证了定理4的结论一和结论二,图 3G验证了定理5,图 3H和图 3I分别验证了定理6的结论一和结论二,图 3J验证了定理7。表明,各混交度的关系取决于空间结构单元的树种数、最近邻木的树种数和成对相邻木非同种的个数以及生物多样性Simpson指数。当空间结构单元树种隔离关系一定时,如果树种多,分布均匀,生物多样性Simpson指数高,则全混交度就大;反之,全混交度就小。树种多样性混交度和物种空间状态之间的关系则服从定理2。显然,全混交度增加考虑了最近邻木相互之间的树种隔离关系,并引入生物多样性Simpson指数,它可以分辨树种空间隔离的细微差别,从而使得树种多样性混交度和物种空间状态之间从2种简单关系(定理2)拓展到3种混交度之间复杂多样的关系,表明全混交度明显优于其他混交度,是描述空间结构单元的树种空间隔离程度的理想指数。
3 结论1) 混交度应全面考虑树种多样性和树种空间隔离关系2个方面。现有混交度虽然考虑了对象木与最近邻木之间的树种隔离关系,但对树种多样性以及最近邻木相互之间的树种隔离关系表达不完整。全混交度增加考虑了最近邻木相互之间的树种隔离关系,并引入Simpson指数,从而提高了对不同混交结构的识别能力,可以对空间结构单元的树种多样性和树种空间隔离关系进行准确表达。
2) 树种多样性混交度、物种空间状态和全混交度都是对简单混交度的改进,因此,各混交度既有区别又有联系。简单混交度≥树种多样性混交度、物种空间状态或全混交度。树种多样性混交度与物种空间状态之间呈分离的线性关系系,即当空间结构单元的树种数=最近邻木的树种数时,树种多样性混交度 > 物种空间状态,当空间结构单元的树种数≠最近邻木的树种数时,树种多样性混交度 < 物种空间状态。全混交度、树种多样性混交度和物种空间状态之间则存在多种复杂关系。
3) 对各混交度进行理论分析和实例应用比较,结果表明:全混交度具有最高的树种隔离程度分辨能力,是较理想的混交度指数。
4) 通常,按照是否考虑林分中树木的位置,把林分结构多样性指标分为与距离有关和与距离无关2类(雷相东等,2002)。事实上,这恰好说明现有林分结构多样性指标对林分结构的描述具有片面性。在全混交度的研究过程中发现,仅从与距离有关的树种相互隔离关系定义混交度,很难区分不同的混交结构单元,但结合与距离无关的生物多样性Simpson指数,问题就解决了。这一研究思路对其他类似研究具有启发性。
| [] | 惠刚盈, 胡艳波, 赵中华. 2008. 基于相邻木关系的树种分隔程度空间测度方法. 北京林业大学学报, 30(4): 131–134. |
| [] | 雷相东, 唐守正. 2002. 林分结构多样性指标研究综述. 林业科学, 38(3): 140–146. DOI:10.11707/j.1001-7488.20020325 |
| [] | 马克平. 1993. 试论生物多样性的概念. 生物多样性, 1(1): 20–22. DOI:10.17520/biods.1993005 |
| [] | 汤孟平, 唐守正, 雷相东, 等. 2004. 两种混交度的比较分析. 林业资源管理(4): 25–27. |
| [] | 汤孟平, 周国模, 施拥军, 等. 2007. 不同地形条件下群落物种多样性与胸高断面积的差异分析. 林业科学, 43(6): 27–31. |
| [] | 汤孟平, 周国模, 陈永刚, 等. 2009. 基于Voronoi图的天目山常绿阔叶林混交度. 林业科学, 45(6): 1–5. DOI:10.11707/j.1001-7488.20090601 |
| [] | 张金屯. 1995. 植被数量生态学方法. 北京, 中国科学技术出版社: 87-89. |
| [] | Gadow von K, Fueldner K. 1992. ZurMethodik der Bestandesbes-chreibung. Vortrag Anlaesslich der Jahrestagung der AG Forsteinrichtung in Klieken b Dessau. |
| [] | Hui G Y, Zhao X H, Zhao Z H, et al. 2011. Evaluating tree species spatial diversity based on neighborhood relationships. Forest Science, 57(4): 292–300. |
| [] | Lexerød N L, Eid T. 2006. An evaluation of different diameter diversity indices based on criteria related to forest management planning. Forest Ecology and Management, 222(1/3): 17–28. |
| [] | Onaindiaa M, Domingueza I, Albizu I. 2004. Vegetation diversity and vertical structure as indicators of forest disturbance. Forest Ecology and Management, 195(3): 341–354. DOI:10.1016/j.foreco.2004.02.059 |
| [] | Pielou E C. 1961. Segregation and symmetry in two-species populations as studied by nearest neighbour relations. Ecology, 49(2): 255–269. DOI:10.2307/2257260 |
| [] | Shimatani K. 2001. Multivariate point processes and spatial variation of species diversity. Forest Ecology and Management, 142(1/3): 215–229. |
| [] | Simpson E H. 1949. Measurement of diversity. Nature, 163: 688. DOI:10.1038/163688a0 |