 2012, Vol. 48
2012, Vol. 48文章信息
- 唐艳龙, 杨忠岐, 王小艺, 唐桦, 姜静, 魏可, 吕军
- Tang Yanlong, Yang Zhongqi, Wang Xiaoyi, Tang Hua, Jiang Jing, Wei Ke, Lü Jun
- 释放花绒寄甲成虫和卵防治栗山天牛
- Biocontrol of Oak Longhorn Beetle, Massicus raddei by Releasing Parasitoid Dastarcus helophoroides (Coleoptera:Bothrideridae) Adults and Eggs
- 林业科学, 2012, 48(7): 186-191.
- Scientia Silvae Sinicae, 2012, 48(7): 186-191.
-
文章历史
- 收稿日期:2012-02-27
- 修回日期:2012-05-14
-
作者相关文章



2. 辽宁宽甸县森林病虫害防治检疫站 宽甸 118200
2. Kuandian Forest Pest & Disease Control Station of Liaoning Province Kuangdian 118200
花绒寄甲(Dastarcus helophoroides) (鞘翅目Coleoptera寄甲科Bothrideridae)是迄今发现的寄生天牛类害虫最主要的寄生性天敌昆虫。经研究,虽然寄生不同天牛种类的花绒寄甲根据形态特征目前认为是同一种,但在生物学特性上表现为很强的寄主专化性,即形成了不同天牛种的花绒寄甲生物型(biotypes)(Wei et al., 2009)。因此,在利用其开展天牛生物防治时,必须在自然界找到自然寄生该种天牛的花绒寄甲生物型,才能取得良好的防治效果。目前已将不同的生物型的花绒寄甲应用于防治云斑天牛(Batocera horsfieldi)、锈色粒肩天牛(Apriona swainsoni)、光肩星天牛(Anoplophora glabripennis)、松褐天牛(Monochamus alternatus)等中大型个体的天牛上(杨忠岐,2004; 张翌楠,2006; 李孟楼等,2009; 李建庆等,2009; 卢希平等,2011; Ogura et al., 1999; Miura et al., 2003; Wei et al., 2009)。杨忠岐(2004)、高峻崇等(2003)在吉林省调查栗山天牛天敌时发现了自然寄生栗山天牛幼虫和蛹的花绒寄甲,其在吉林省梅河口市鸡冠山林场的自然寄生率为10%左右,但在其他林区寄生率很低,自然分布呈团块型。随后,经过多年研究,解决了花绒寄甲的人工大量繁殖问题,并先后在中国林科院、吉林省梅河口市和辽宁省宽甸县森林病虫害防治站建立了栗山天牛花绒寄甲繁育室袁繁育出了大量花绒寄甲成虫和卵袁在生产防治中应用于大面积释放防治栗山天牛袁防治面积达11万hm2(魏建荣等,2007; 2009; 唐艳龙等,2010)。在山东省进行的生物防治试验结果显示:释放天敌后栗山天牛的虫口减退率达到88%(魏建荣等,2009),表明花绒寄甲对栗山天牛具有良好的防治效果。为进一步明确释放花绒寄甲成虫和卵防治栗山天牛的最佳虫期和时期及释放数量比例,从2004年起笔者在吉林、辽宁省开展了释放花绒寄甲防治栗山天牛试验,并调查了花绒寄甲对栗山天牛的持续控制效果,现将结果报道如下。
1 材料与方法 1.1 室内花绒寄甲寄生试验2011年5月,栗山天牛处于老龄幼虫期,在辽宁省宽甸县大西岔镇(40°44.846′ N,125°11.625′ E)受栗山天牛危害严重的辽东栎(Quercus wutaishanica)林地伐树解剖采集栗山天牛幼虫,选择健康的幼虫用作试虫。依据林间天牛幼虫所在的位置,将试虫接于长40 cm、直径15 cm的辽东栎木段内。每个木段接幼虫10头,待幼虫完全蛀入木段并开始往外排木屑和虫粪之后,将木段置于长50 cm、直径20 cm的有底无盖玻璃缸内,按2:1、1:1和1:2(寄甲数量:天牛数量)的比例接花绒寄甲成虫于木段外,并用纱布盖住玻璃缸口,然后用皮筋套紧。每个比例重复3次,每个重复处理45头天牛幼虫。玻璃缸置于室内室温条件下,采用自然光照饲养。1个月之后,劈开上述木段,检查天牛幼虫被寄生情况。观察到栗山天牛幼虫上有花绒寄甲幼虫寄生或坑道中有花绒寄甲茧,则视为该天牛被寄生,统计寄生率。
分别在栗山天牛幼虫的1~6龄幼虫期(1龄幼虫期2008年9月,2龄幼虫期2009年6月,3龄幼虫期2009年7月,4龄幼虫期2009年9月,5龄幼虫期2010年7月,6龄幼虫期2011年6月) (王小艺等,2012),按上述方法采集和处理栗山天牛幼虫,按1:2 (天牛数量:寄甲数量)的比例释放花绒寄甲成虫,同时另设一处理,按1:100 (天牛数量:寄甲卵粒数)的比例接花绒寄甲卵粒于木段外。每次设3个重复,1~5龄幼虫期每个重复处理30头天牛幼虫,6龄幼虫期每个重复处理45头天牛幼虫。1个月之后,按上述方法统计寄生率。同时记录栗山天牛被寄生后子代花绒寄甲幼虫或茧的数量,并将幼虫或茧在室内饲养至成虫羽化后计数。按照公式: F2/F1=子代花绒寄甲成虫数量/释放花绒寄甲成虫数量来计算释放成虫寄生栗山天牛后子代花绒寄甲增加的倍数。按照公式:M=F2/2×P/P1计算释放花绒寄甲卵寄生栗山天牛后子代花绒寄甲产卵量与释放量的比值。式中,M为花绒寄甲卵寄生栗山天牛后子代花绒寄甲产卵量与释放量的比值,F2为子代花绒寄甲成虫数量,P为花绒寄甲雌虫平均年产卵量(秦锡祥等,1988),P1为释放花绒寄甲卵粒数遥花绒寄甲成虫雌雄性比按1:1计算(魏建荣等,2007)。
1.2 林间防治试验于栗山天牛5龄和6龄幼虫期,在辽宁省宽甸县大西岔镇选择受栗山天牛危害较重的栎树作为样株袁将样株伐倒并截成2m长的木段,根据一个栗山天牛幼虫一般只有1个排粪孔的生物学特性,统计样株栗山天牛的排粪孔数,用网眼为1 mm×1 mm的纱网做成笼罩,将木段置于笼罩内,按寄甲数量:天牛数量2:1、1:1和1:2的比例接花绒寄甲成虫于木段上,然后扎紧网口,置于林间。每个比例处理3株。同时在林间选择10株样株不伐倒,统计其2 m以下主干上的排粪孔数,在每个排粪孔处释放花绒寄甲卵100粒。1个月后,劈开上述样株,仔细检查其中的栗山天牛幼虫被寄生情况。天牛幼虫体上有寄生的花绒寄甲幼虫或坑道中有花绒寄甲茧,则该天牛统计为被寄生,计算寄生率。同时统计栗山天牛被寄生后子代花绒寄甲幼虫或茧的数量,并将幼虫或茧在室内饲养至成虫羽化后计数。按上述公式计算子代花绒寄甲增加的倍数。同时在未释放花绒寄甲的林区,选择受害较重的栎树伐倒解剖检查花绒寄甲自然寄生情况,作为对照。
1.3 林间释放花绒寄甲对栗山天牛的寄生率调查2008年5月,栗山天牛处于老龄幼虫期,在上述林区选择受害较重的栎树林作为样地,选择标准株3株,伐倒劈开,统计栗山天牛幼虫的数量,计算平均虫口密度,按天牛幼虫:寄甲成虫1:2的数量比例在林间释放花绒寄甲成虫,释放面积15 hm2。之后在栗山天牛不同龄期(1~6龄),在试验林区砍伐标准株3株,劈开检查栗山天牛被花绒寄甲寄生情况,计算寄生率; 同时在未释放花绒寄甲的林区(相隔2 000 m以上)选择样株3株,伐倒劈开统计栗山天牛数量及花绒寄甲自然寄生数量,作为对照。
1.4 释放花绒寄甲后林间花绒寄甲种群动态调查2008—2011年,在上述释放过花绒寄甲成虫防治栗山天牛的试验林区,设置诱捕器。每隔20 m设置1个,山脊、阳坡和阴坡各设置2排,每排9个,一共设置54个诱捕器。诱捕器用铁丝悬挂在林间距地表 1.5 m高的位置处。所用诱捕器为北京中捷四方生物科技有限公司生产的漏斗形小蠹虫诱捕器。诱捕器内的诱芯添加对花绒寄甲有引诱作用的柠檬烯类引诱剂(Wei et al., 2008)。
2008—2011年每年6—9月,于每月月末检查1次诱捕器,采集诱捕器中诱到的花绒寄甲成虫并计数。
1.5 数据分析用Excel 2003和SPSS 11.0等软件进行数据处理,用SPSS中的One-Way ANOVA进行方差分析,用Duncan氏新复极差法检验,比较各处理之间的差异水平(贾乃光,1993)。
2 结果与分析 2.1 室内接花绒寄甲成虫对栗山天牛的寄生试验在室内试验中,按不同比例接入花绒寄甲成虫对栗山天牛的寄生率差异显著(F=6.319; df=2, 8; P=0.033 4)(图 1)。按天牛幼虫:寄甲成虫1:2的数量比例接入的,寄生率最高达88.15%;按1:1释放比例接入的,寄生率达到80.74%;按2:1接入的,寄生率为70.37%,显著低于释放比例为1:2时的寄生率。

|
图 1 室内接入不同比例花绒寄甲成虫对栗山天牛的寄生率 Fig.1 Introducing D. helophoroides adults with different ratio to control M. raddei in lab 释放比例Releasing ratio:栗山天牛幼虫数:花绒寄甲成虫数量Number of oak longhorn beetle larva:Number of D. Helophoroides adult. |
在室内试验中,按天牛幼虫:寄甲成虫1:2的数量比例,接入花绒寄甲成虫对栗山天牛不同龄期幼虫的寄生率差异极显著(F=517.283; df=5, 17;P < 0.000 1)(图 2)。栗山天牛处于1~3龄幼虫期时,未发现花绒寄甲寄生。栗山天牛幼虫为4龄时,发现花绒寄甲开始寄生,不过寄生率较低,不足5%。栗山天牛到5龄幼虫时,寄生率达到了34.44%。栗山天牛处于老龄幼虫(6龄)和蛹期时,寄生率最高达88.15%,显著高于其他幼虫龄期时接入的寄生效果。

|
图 2 室内接入花绒寄甲成虫对栗山天牛不同龄期幼虫的寄生率 Fig.2 Parasitization rate of M. raddei in different instars by introducing D. helophoroides adults in lab |
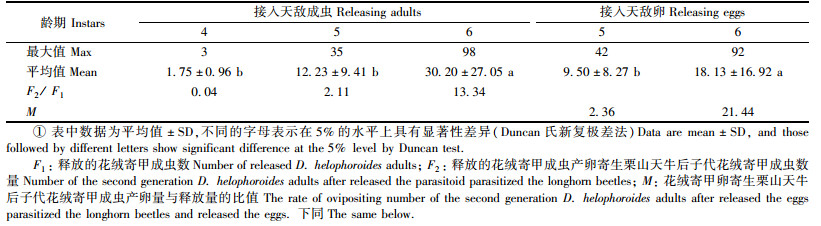
接花绒寄甲成虫后寄生栗山天牛后袁其子代成虫数量差异较大(表 1)。栗山天牛为4龄幼虫时,只有少数天牛被寄生,因而花绒寄甲子代数量亦较少,每头寄主上平均只有1.75头; 栗山天牛为5龄幼虫时,被寄生的天牛明显增多,每头寄主上子代数最多有35头,平均为12.23头,花绒寄甲子代成虫数量是接入量的2.11倍; 栗山天牛处于6龄幼虫期和蛹期时,绝大多数天牛都被寄生,每头寄主上子代数最多达98头(图 3),平均为30.2头,明显高于其他时间子代成虫的数量,子代成虫数量是接入量的13.34倍。试验中发现:即使只1头花绒寄甲寄生栗山天牛幼虫或蛹都会致寄主死亡(图 4)。
|
|

|
图 3 取食完寄主幼虫后作茧化蛹的花绒寄甲幼虫 Fig.3 Cocoons of D. helophoroides which consumed up its host larva after parasitizing |

|
图 4 寄生于栗山天牛蛹上的花绒寄甲幼虫 Fig.4 D. helophoroides larva parasitizing on pupa of oak longhorn beetle |
在室内试验中,按天牛幼虫数量:寄甲卵粒数1:100的比例,接花绒寄甲卵对栗山天牛不同龄期幼虫的寄生率差异极显著(F=629.860,df=5, 17,P < 0.000 1)(图 5)。不同时间接的卵粒孵化率均在90%以上。栗山天牛为1~4龄幼虫时,未发现花绒寄甲寄生; 当栗山天牛为5龄幼虫时,发现天敌开始寄生,但寄生率只有17.78%;当栗山天牛为6龄幼虫时,寄生率明显上升,最高达90%,显著高于其他龄期接入的寄生率。

|
图 5 室内接入花绒寄甲卵对栗山天牛不同龄期幼虫的寄生率 Fig.5 Parasitization rate of M. raddei in different instars by introducing D. helophoroides eggs in lab |
室内接花绒寄甲卵寄生栗山天牛幼虫的子代成虫数量差异较大(表 1)。栗山天牛处于老龄幼虫期和蛹期时,大多数天牛都被寄生,每头寄主上子代数最多达92头(图 3),平均为18.13头,显著高于其他时间接入的子代成虫的数量,按每头雌虫年产280粒卵计算,得出子代卵粒数量是释放量的21.44倍。栗山天牛为5龄幼虫时,天敌的寄生率还较低,每头寄主上子代数最多有42头(图 3),平均为9.5头,子代卵粒数量是接入量的2.36倍。与接入成虫不同的是,栗山天牛为4龄幼虫时,接入花绒寄甲卵未发现幼虫被寄生。
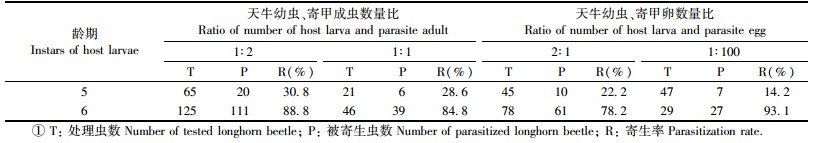
2.3 林间释放花绒寄甲成虫和卵对栗山天牛的寄生试验在林间,栗山天牛处于中老龄幼虫期和蛹期释放花绒寄甲成虫和卵防治栗山天牛效果良好(表 2)。其中以老龄幼虫和蛹期释放效果最佳,按天牛幼虫数量:寄甲数量1:2的比例释放成虫时,其对天牛的寄生率高达88.8%,按1:1的比例释放时,寄生率为84.8%,按2:1的比例释放时,寄生率略低,为78.2%。在栗山天牛5龄幼虫期时释放,寄生率为30%左右。栗山天牛处于老龄幼虫期和蛹期时按1:100的比例释放花绒寄甲卵时,卵粒的孵化率均在80%左右,其对栗山天牛的寄生率比释放成虫还高,达93.1%。对照林区只发现少数栗山天牛被花绒寄甲寄生,寄生率不足5%。
|
|
林间释放花绒寄甲成虫寄生栗山天牛后的子代成虫数量差异较大(表 3)。栗山天牛处于老龄幼虫期和蛹期时,大多数天牛都被寄生,子代数最多达每窝92头,平均为每窝27.97头,明显高于其他时期子代成虫的数量,是释放量的17.62倍。释放花绒寄甲卵寄生天牛后的子代成虫数量差异亦较大(表 3),在栗山天牛老龄幼虫期和蛹期,子代成虫数量平均为每窝10头,按每头雌虫年产卵280粒计算,得出子代卵粒数量是释放量的13.52倍。
|
|

2008年在试验林间释放花绒寄甲后,其对栗山天牛的寄生率变化呈峰型变化(表 4)。栗山天牛处于老龄幼虫期和蛹期时,寄生率最高达85.46%,栗山天牛处于1~3龄幼龄幼虫期时,未发现花绒寄甲寄生; 栗山天牛为4龄幼虫时,发现花绒寄甲开始寄生,但寄生率较低,只有3.25%;到栗山天牛为5龄幼虫时,寄生率增加到22.50%;栗山天牛处于老龄幼虫期和蛹期时,寄生率增加到75.80%。表明花绒寄甲对栗山天牛有较强的持续控制作用。对照林区只发现少数栗山天牛被花绒寄甲寄生,寄生率不足5%。
|
|

林间不同时间诱捕到的花绒寄甲成虫数量变化较大(图 6),每年的7—8月诱捕到的成虫数量较多,而6月和9月诱捕到的成虫数量较少。2008年7月诱捕到的成虫数量最多,为180头,这与花绒寄甲寄生栗山天牛老龄幼虫和蛹后成虫数量大量增加有关。而在不适合花绒寄甲寄生的栗山天牛中幼龄幼虫期,花绒寄甲成虫的数量有所下降,2009年7月和2010年7月,分别诱捕到成虫138头和95头。在寄生下一代栗山天牛老龄幼虫和蛹后,花绒寄甲的种群数量又上升到较高水平。2011年7月诱捕到成虫177头,只比2008年7月诱捕到的成虫数量略少。

|
图 6 林间不同时间诱捕到的花绒寄甲成虫数量 Fig.6 Number of D. helophoroides adults trapped in field in different time |
分布在山脊、阳坡和阴坡的花绒寄甲成虫数量差异达到了极显著水平(F=22.523; df=2, 11; P=0.000 3)(图 7)。分布在山脊的成虫数量最多,平均每年诱捕到176头。阳坡次之,平均每年诱捕到90头,阴坡最少,平均每年诱捕到34头,而且主要是分布在阴坡的上坡位。

|
图 7 花绒寄甲成虫的空间分布 Fig.7 Distribution of D. helophoroides adult |
栗山天牛在危害严重的东北地区3年1代,而且发育比较整齐(侯义等,2000)。在栗山天牛中老龄幼虫期和蛹期,室内和林间释放花绒寄甲成虫和卵来防治栗山天牛效果良好。释放成虫的试验中,花绒寄甲对栗山天牛的寄生率达88%,释放卵的试验中,寄生率达到了90%,花绒寄甲子代数量增加了20多倍,即使是在林间不采取保护措施的情况下释放天敌,其寄生率也达到了85%。因此,利用花绒寄甲来防治栗山天牛,把握好释放时间是成功的关键。本研究表明:栗山天牛中老龄幼虫期和蛹期寄生率最高。因此,栗山天牛中老龄幼虫期和蛹期是释放花绒寄甲成虫防治栗山天牛的最佳时期。虽然本试验中释放的天敌数量较多,在生产实际中需要较大的投入,但是一次大量释放花绒寄甲之后,可在短期内大量寄生了害虫,得到了显著的防治效果,而且由于大大增加了林间花绒寄甲的种群数量,因此在栗山天牛下一个适合花绒寄甲寄生的时期出现时,其在林间的种群数量仍然较大,从而能够保持对栗山天牛的长期的持续控制作用。本试验林区的天牛虫口密度在经过花绒寄甲2个世代的寄生之后,由每株89头下降到不足每株5头,而未释放天敌的林区,虫口密度仍呈增加趋势。
花绒寄甲不仅能寄生栗山天牛,对云斑天牛、松褐天牛、光肩星天牛和锈色粒肩天牛等都能寄生,表明其是一种多寄主型天敌昆虫,但表现为明显的寄主专化性,因而分化形成了不同的花绒寄甲寄主生物型(杨忠岐,2004; 张翌楠,2006; 李孟楼等,2009; 李建庆等,2009; 卢希平等,2011; Ogura et al., 1999; Miura et al., 2003; Wei et al., 2009)。花绒寄甲在环境多变的条件下能够保持较高的种群数量,从而对害虫起到长期控制的效果。由于寄生栗山天牛的花绒寄甲经笔者研究只在4龄以上的寄主幼虫期寄生,而表现为在幼龄幼虫期不寄生的习性。为了达到良好的防治效果,笔者试验利用栗山天牛1~3龄幼龄幼虫期释放白蜡吉丁肿腿蜂(Sclerodermus pupariae)进行防治,也取得了良好的防治效果(王小艺等,2010; 唐艳龙等,2011)。通过在栗山天牛不同发育阶段利用这2种天敌,相互配合使用,实现了对栗山天牛的有效控制,防治效果显著。
| [] | 高峻崇, 山广茂, 赵海滨, 等. 2003. 吉林省首次发现捕食栗山天牛的天敌—花绒坚甲. 吉林林业科技, 32(1): 45–47. |
| [] | 侯义, 季长龙, 高纯, 等. 2000. 栗山天牛生物学特性及防治技术研究. 辽宁林业科技, 19(5): 15–18. |
| [] | 贾乃光. 1993. 数理统计. 2版. 北京, 中国林业出版社. |
| [] | 李建庆, 杨忠岐, 张雅林, 等. 2009. 利用花绒寄甲防治杨树云斑天牛的研究. 林业科学, 45(9): 94–100. DOI:10.11707/j.1001-7488.20090916 |
| [] | 李孟楼, 李有忠, 雷琼, 等. 2009. 释放花绒寄甲卵对光肩星天牛幼虫的防治效果. 林业科学, 45(4): 78–82. DOI:10.11707/j.1001-7488.20090413 |
| [] | 卢希平, 杨忠岐, 孙绪艮, 等. 2011. 利用花绒寄甲防治锈色粒肩天牛. 林业科学, 47(10): 116–121. DOI:10.11707/j.1001-7488.20111018 |
| [] | 秦锡祥, 高瑞桐. 1988. 花绒寄甲生物学特性及其应用研究. 昆虫知识, 25(2): 109–112. |
| [] | 唐艳龙, 杨忠岐, 魏建荣, 等. 2010. 栗山天牛研究进展. 中国森林病虫, 29(3): 12–17. |
| [] | 唐艳龙, 杨忠岐, 姜静, 等. 2011. 栗山天牛幼虫和蛹在辽东栎树干的分布规律. 林业科学, 47(4): 117–123. |
| [] | 唐艳龙, 王小艺, 杨忠岐, 等. 2012. 栗山天牛幼虫天敌白蜡吉丁肿腿蜂的转主寄主研究. 昆虫学报, 55(1): 55–62. |
| [] | 王小艺, 杨忠岐, 唐艳龙, 等. 2010. 白蜡吉丁肿腿蜂对栗山天牛低龄幼虫的寄生作用. 昆虫学报, 53(6): 675–682. |
| [] | 王小艺, 杨忠岐, 唐艳龙, 等. 2012. 栗山天牛幼虫的龄数和龄期测定. 昆虫学报, 55(5): 574–583. |
| [] | 魏建荣, 杨忠岐, 马建海, 等. 2007. 花绒寄甲研究进展. 中国森林病虫, 26(3): 23–25. |
| [] | 魏建荣, 杨忠岐, 王平彦, 等. 2009. 利用花绒寄甲控制栗山天牛林间试验效果. 中国生物防治, 25(3): 285–287. |
| [] | 杨忠岐. 2004. 利用天敌昆虫控制我国重大林木害虫研究进展. 中国生物防治, 20(4): 221–227. |
| [] | 张翌楠. 2006. 松褐天牛的天敌昆虫调查及生物防治技术研究. 中国林业科学研究院博士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-82201-2006152450.htm |
| [] | Miura K, Abe T, Nakashima Y, et al. 2003. Field release of parasitoid Dastarcus helophoroides (Fairmaire) (Coleoptera:Bothrideridae) on pine logs infested with Monochamus alternatus Hope (Coleoptera:Cerambycidae) and their dispersal. Journal Japanese Forestry Society, 85(1): 12–17. |
| [] | Ogura N, Tabata K, Wang W. 1999. Rearing of the colydiid beetle predator, Dastarcus helophoroides, on artificial diet. Biocontrol, 44(3): 291–299. DOI:10.1023/A:1009936609401 |
| [] | Wei J R, Yang Z Q, Hao H L, et al. 2008. (R)-(+)-limonene, kairomone for Dastarcus helophoroides, a natural enemy of longhorned beetles. Agricultural and Forest Entomology, 10(1): 1–8. |
| [] | Wei J R, Yang Z Q, Poland T M, et al. 2009. Evaluation of the behavioral response and parasitism of Dastarcus helophoroides among different Cerambycidae hosts. Biocontrol, 54(6): 733–742. DOI:10.1007/s10526-009-9224-y |