 2012, Vol. 48
2012, Vol. 48文章信息
- 刘奎伟, 孔祥波, 张真, 周成刚, 柴景峰
- Liu Kuiwei, Kong Xiangbo, Zhang Zhen, Zhou Chenggang, Chai Jingfeng
- 共轭二烯性信息素在2种载体上的释放速率和异构化效果
- Release Rates and Isomerization Effects of Conjugated Dienes Sex Pheromone from Two Types of Dispensers
- 林业科学, 2012, 48(7): 72-78.
- Scientia Silvae Sinicae, 2012, 48(7): 72-78.
-
文章历史
- 收稿日期:2012-02-22
- 修回日期:2012-05-16
-
作者相关文章



2. 山东农业大学植物保护学院 泰安 271018;
3. 内蒙古克什克腾旗桦木沟林场 克什克腾 025350
2. College of Plant Protection, Shandong Agricultural University Tai'an 271018;
3. Huashugou Forest Farm of Keshiketeng Banner of Inner Mongolia Keshiketeng 025350
鳞翅目蛾类昆虫性信息素成分根据其化学结构特征分为2大类,一类是具有10~18个碳原子的直链烯烃,碳链末端具有醇、醛或酯类官能团,碳链上具有1~3个双键,有2个以上双键时多以共轭形式存在;另一类是具有17~23个碳原子的直链烯烃,碳链上1~4个顺式双键由亚甲基分开,且具有0,1或者2个环氧官能团(Millar, 2000)。利用性信息素防控农林害虫时要求其成分在害虫发生期内能够稳定持续释放,以保持仿生诱芯的持久生物活性,这对性信息素释放载体的选择提出了较高的要求。目前常用的性信息素释放载体有橡胶塞(Vrkoč et al., 1988; Kong et al., 2011)、聚乙烯管(Hodges et al., 2004; Cork et al., 2001)、聚氯乙烯树脂(Cork et al.,2008)、空心纤维(由聚对苯二甲酸酯构成)(Weatherston et al., 1985)和塑料薄片等。不同的释放载体对不同的性信息素成分有不同的释放特性,而适宜的释放载体不仅能保证有效成分的稳定释放,还应保护其在载体中不被氧化降解或受载体诱导发生异构化(Ideses et al., 1982; Brown et al., 1986)。例如,共轭二烯类性信息素成分在含硫橡胶塞载体上容易发生异构化(Vrkoč et al., 1988; Brown et al., 1986),而硅橡胶材料对此影响较小(荆慧荣等,1996),但持效期短。所以橡胶载体应具有一定的交联度且不含易与性信息素成分发生作用的成分,以保证性信息素成分均匀持续释放。通常认为酚醛树脂处理或者过氧化物处理的卤代丁基橡胶载体是适宜的性信息素释放载体(Brown et al., 1986)。另外,在配制仿生诱芯时加入紫外光吸附剂(2-羟基-4甲氧基二苯甲酮)和抗氧化剂(BHT或BHA)(Ideses et al., 1988)都是保护有效成分避免光氧化非常必要的措施。
研究载体中挥发性性信息素成分释放速率的方法包括动态顶空吸附法(Weatherston et al., 1981)、静态瓶壁吸附法(Vrkoč et al., 1988)、残留量测定法(Cork et al., 2008; Alfaro et al., 2009)和称重法(Rothschild, 1979)等,不同的测量方法各有优缺点。对于动态顶空吸附法,挥发性信息素成分的收集方法包括冷阱、溶剂和吸附剂(如活性炭、Porapak Q和Tenax GR)法。松毛虫(Dendrolimus spp.)性信息素化学成分一般由顺5, 反7-或者反5, 顺7-十二碳二烯醇、醛和乙酸酯构成(Kong et al., 2001; 2007a; 2007b; 2011),在载体中容易发生异构化而丧失生物活性。本文采用Porapak Q吸附剂收集复合橡胶载体和聚乙烯管2种载体上6种松毛虫性信息素成分的释放量,研究性信息素成分在不同载体上的释放速率及异构化程度,以此指导仿生诱芯的配制,提高害虫种群监测效果。
1 材料与方法 1.1 性信息素成分、释放载体和吸附剂顺5, 反7-十二碳二烯醛(Z5, E7-12: Ald),反5, 顺7-十二碳二烯醛(E5, Z7-12: Ald),顺5, 反7-十二碳二烯醇(Z5, E7-12: OH),反5, 顺7-十二碳二烯醇(E5, Z7-12: OH),顺5, 反7-十二碳二烯乙酸酯(Z5, E7-12: OAc)和反5, 顺7-十二碳二烯乙酸酯(E5, Z7-12: OAc)6种松毛虫性信息素成分均购自Chemtech B.V.公司(Amsterdam, The Netherlands)。灰色袖口式医用丁基复合胶塞购自West Pharmaceutical Services, Inc.(Phoenixville, PA, USA),聚乙烯管(polyethylene tube 1 mL;32 mm long × 8 mm OD × 6 mm ID)购自Kartell Co.,这2种载体使用前均用无水乙醇浸泡24 h。吸附剂Porapak Q(50~80目)购自CNW Technologies,以每采样管150 mg填充到玻璃采样管中(Chrompack, Φ 0.6 mm × 160 mm),使用前在200 ℃高温条件下老化2 h。
1.2 性信息素诱芯的制备本试验将测试的性信息素分为2组,第1组含有E5, Z7-12: Ald、E5, Z7-12: OH和E5, Z7-12: OAc;第2组含有Z5, E7-12: Ald、Z5, E7-12: OH和Z5, E7-12: OAc。每组3个成分各1 mg混合均匀,滴加到聚乙烯管载体和复合橡胶载体,每组在每种载体上重复3次。2种载体中分别加入0.5 mg抗氧化剂BHT。溶剂挥发后将聚乙烯管载体管口密封,复合橡胶载体以开口的形式用铁丝悬挂于风洞的气流中。
1.3 2种载体上挥发性性信息素成分的采集在风洞实验室内进行气味的采集,风速20 cm·s-1,温度23~27 ℃,相对湿度50%~80%。大气采样仪(QC-1型,北京市劳动保护科学研究所)进气口端以特氟隆管连接采样管,以100 mL·min-1的气体流速采样20 min。点样后第2天采集1次挥发性性信息素成分,然后每2周采集1次。采集的样品管立即用色谱纯的正己烷洗脱,洗脱液(4 mL)在微弱的氮气流下浓缩至300 μL备用。
1.4 色谱分析采用7683B series自动进样器和7890A-GC(Agilent, Palo Alto, CA, USA)色谱仪分析正己烷洗脱液。氮气作载气,无分流进样(0.75 min后开阀),检测器为火焰离子化检测器(FID)。分析条件是:DB-5MS毛细管色谱柱(30 m × 0.25 mm × 0.25 μm),60 ℃保持0.75 min,以6 ℃·min-1升温至250 ℃,保持15 min;进样器温度230 ℃,检测器温度250 ℃,柱头压力10 Pa。用Agilent Chemstation软件进行数据采集分析。通过色谱保留时间和质谱定性,采用外标法定量。分别配制6种共轭二烯性信息素成分5个浓度梯度(0.1,0.5,2.5,12.5和62.5 ng·μL-1),进行7890A-GC分析(条件同上),建立浓度和峰面积的标准曲线,利用标准曲线进行定量。
1.5 性信息素成分残留量的测定最后一次释放速率采样完成后,将2种载体剪碎,分别浸泡在1.2 mL的二氯甲烷中3天,然后取1 μL进GC分析,计算异构体的比例和残留量。色谱分析方法同上。
1.6 数据分析表中数据均以Mean ± SE表示。数据经lg(x+1) 转化后进行单因素方差分析(ANOVA),若F值差异显著,再进行SNK或Duncan多重比较不同处理平均数间的差异显著性(SPSS 13.0)。聚乙烯管载体和复合橡胶载体中不同样品的异构化率和剩余量进行Independent Samples T-tests显著性检验。异构化率用公式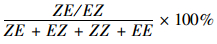
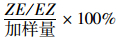
用DB-5MS毛细管色谱柱分析共轭二烯醇、醛和酯类性信息素成分在聚乙烯管载体和复合橡胶载体中不同时间的释放速率(图 1)。对于聚乙烯管载体(PE, 吸附剂Porapak Q),顺5, 反7-和反5, 顺7-共轭二烯醇、醛和酯类性信息素成分具有相似的释放模式(图 1a):第2~15天是性信息素释放的初级阶段,即点样后的缓慢平稳释放阶段;从第16天开始释放速率一直上升,第29天达到最大值(Z5, E7-12: Ald是第43天达到释放最大值),然后又缓慢下降,第57天后6种性信息素成分处在较稳定的低释放速率状态。在释放高峰期内,反5, 顺7-共轭二烯醇、醛和酯类性信息素成分的释放速率快于相应的顺5, 反7-共轭二烯类性信息素成分,且醛和醇类性信息素成分的释放速率大于酯类性信息素成分。6种性信息素成分在第15天(F(5,12)=1.42, P=0.286 > 0.05) 和第29天(F(5,12)=2.15, P=0.129 > 0.05) 的释放速率差异不显著;第43天的释放速率间差异显著(F(5,12)=11.92, P < 0.001),多重比较(SNK,α=0.05) 发现E5, Z7-12: Ald,E5, Z7-12: OH和Z5, E7-12: Ald的释放速率显著大于Z5, E7-12: OH,Z5, E7-12: OAc和E5, Z7-12: OAc的释放速率。第57天Z5, E7-12: OH的释放速率显著低于E5, Z7-12: OH的释放速率(F(5,12)=3.89, P=0.025 < 0.05),其他成分间差异不显著。

|
图 1 顺5, 反7-和反5, 顺7-十二碳二烯醇、醛和乙酸酯性信息素在聚乙烯管载体(a)和复合橡胶载体(b)上释放速率 Fig.1 Dynamics of release rates of Z5, E7-and E5, Z7-dodecadien-1-ol, and their corresponding acetates and aldehydes derivatives in PE (a) and rubber (b) dispensers |
对6种性信息素成分在不同时间的释放速率分别进行方差分析,发现其在第2,57,71和81天4个时间段内差异不显著。上述4个时间段与第29天和第43天的释放速率差异显著。例如,E5, Z7-12: Ald的释放速率在7个分析时间段间差异极显著(F(6, 14)=42.98, P < 0.001),多重比较分析(SNK, α=0.05) 发现:第29天和43天的平均释放速率为(32.37 ± 20.89)μg·d-1和(19.67 ± 4.35)μg·d-1,显著高于其他测试时间,第15天的平均释放速率是(10.35 ± 3.91)μg·d-1,与其他时间段也差异显著。在第2,57,71和81天的平均释放速率分别是(0.35 ± 0.23),(1.25 ± 0.65),(1.58 ± 0.29) 和(0.32±0.22)μg·d-1,释放速率间差异不显著。在聚乙烯管载体上6种性信息素成分释放高峰期内的比例变化:(Z5, E7-12: Ald):(Z5, E7-12: OH):(Z5, E7-12: OAc)分别为100:92:72(第15天)、100:83:67(第29天)和100:35:33(第43天)。(E5, Z7-12: Ald):(E5, Z7-12: OH):(E5, Z7-12: OAc)分别为100:156:82(第15天)、100:104:42(第29天)和100:80:45(第43天)。
对于复合橡胶载体(Rubber, 吸附剂Porapak Q),顺5, 反7-和反5, 顺7-共轭二烯醇、醛和酯类性信息素成分的释放模式与聚乙烯管载体完全不同(图 1b):点样后第2天测试时释放量最大,第2~15天是释放最快的时期,第15~43天释放速率逐渐降低,第44~81天释放速率维持在低释放状态。复合橡胶载体中6种性信息素成在第15天的释放速率存在显著差异(F(5,12)=8.13, P=0.01)。经Duncan多重比较发现:Z5, E7-12: OH(9.09±4.88)μg·d-1的释放速率显著高于其他5种性信息素成分,其次为E5, Z7-12: OH(3.41±1.36)μg·d-1,其余4种信息素成分的释放速率间差异不显著。在释放转折点第29天,Z5, E7-12: OH(2.31±1.67)μg·d-1与E5, Z7-12: Ald(0.18±0.01)μg·d-1的释放速率差异显著(F(5,12)=3.64, P=0.03)。
对6种性信息素成分在不同测试时间内的释放速率进行方差分析,发现Z5, E7-12: Ald、Z5, E7-和E5, Z7-12: OH、Z5, E7-和E5, Z7-12: OAc存在显著性差异。上述5种性信息素成分的释放速率在7个测试时间段分别进行多重比较分析,发现在第57,71和81天的释放速率间差异不显著,第2天和第15天的释放速率与其他时间段存在显著性差异。例如,E5, Z7-12: OH在第29,43,57,71和81天处释放速率分别为(0.34±0.01),(0.83±0.28),(0.33±0.11),(0.58±0.41) 和(0.18±0.07)μg·d-1,相互间差异不显著,但是第2天和第15天的释放速率分别为(8.22±6.65) 和(3.41±1.36)μg·d-1,与其他5个测试时间段间存在显著差异(F(6,14)=9.06, P < 0.001)。Z5, E7-12:OH在第43,57,71和81天的释放速率分别为1.04±0.40,0.81±0.13,0.30±0.20和(0.31±0.23)μg·d-1,差异不显著;第2天和第15天的释放速率分别为(8.41±6.96) 和(9.09±4.88)μg·d-1,与其他5个测试时间段间存在显著差异(F(6,14)=5.88, P =0.03 < 0.05);第29天释放速率为(2.31±1.67)μg·d-1,处于中间水平。在复合橡胶载体上,6种性信息素成分释放高峰期内的比例变化:(Z5, E7-12: Ald):(Z5, E7-12: OH):(Z5, E7-12: OAc)分别为100:191:29(第2天)、100:648:68(第15天)和100:189:19(第29天)。(E5, Z7-12: Ald):(E5, Z7-12: OH):(E5, Z7-12: OAc)分别为100:1 275:317(第2天)、100:450:205(第15天)和100:188:302(第29天)。
在释放高峰期内6种性信息素成分在不同的载体中释放速率不同,信息素成分在聚乙烯管载体上的释放速率大于复合橡胶载体上的释放速率(图 1)。对聚乙烯管载体第29天的释放速率与复合橡胶载体第15天的释放速率比较分析发现,除Z5, E7-12: OH(F(1, 4)=0.57, P=0.49) 外,E5, Z7-12: Ald(F(1, 4)=41.76, P=0.001)、E5, Z7-12: OH(F(1, 4)=19.57, P=0.011)、E5, Z7-12: OAc(F(1, 4)=31.99, P=0.005)、Z5, E7-12: Ald(F(1, 4)=24.79, P=0.008) 和Z5, E7-12: OAc(F(1, 4)=99.78, P=0.001)5种性信息素成分在2种载体上的释放速率差异显著。
2.2 性信息素成分在2种载体上的剩余量和异构化分析对相同性信息素成分在不同载体上的剩余量进行Independent Samples T-tests分析,除Z5, E7-12: OH(t=-2.59,df=4,P=0.60) 外,性信息素成分在复合橡胶载体中的剩余量与聚乙烯管载体中的剩余量差异极显著(表 1)。例如,Z5, E7-12: OAc在橡胶载体和聚乙烯管载体上的剩余量分别是初始点样量的19.17%和2.96%。
|
|

对6种性信息素成分在2种载体上的异构化率进行Independent Samples T-tests分析,Z5, E7-12: Ald和Z5, E7-12: OAc 2种性信息素成分的异构化率在2种载体上差异不显著,其他4种性信息素成分E5, Z7-12: Ald(t=-3.80,df=4,P=0.019)、E5, Z7-12: OH(t=21.62,df=4,P=0.001)、E5, Z7-12: OAc(t=3.781,df=4,P=0.019) 和Z5, E7-12: OH(t=-9.418,df=4,P=0.001) 的异构化率差异显著。除载体中E5, Z7-12: Ald(聚乙烯管载体t=-1.218,df=4,P=0.29;复合橡胶载体t=0.72,df=4,P=5.11) 外,其他性信息素成分在2种释放载体中都是E5, E7异构体成分的含量显著大于Z5, Z7异构体成分的含量。另外,发现2种不同载体中醇的异构化形式与其他性信息素成分不一样,即ZE(EZ) > EE > ZZ,2个双键都发生异构化的概率大于单个双键发生异构化的概率。聚乙烯管载体中6种性信息素成分的异构化率存在显著差异(F(5, 12)=120.40,P < 0.001),其中E5, Z7-12: OH的异构化率92.48%显著高于其他性信息素成分的异构化率,Z5, E7-12: Ald的异构化率25.38%也显著高于其他4种成分的异构化率。异构化率最小的是E5, Z7-12: Ald和Z5, E7-12: OH。复合橡胶载体中6种性信息素成分的异构化率也存在显著性差异(F(5,12)=98.31,P < 0.001),E5, Z7-12: OH和Z5, E7-12: OH的异构化率显著大于其他4种性信息素成分的异构化率。
3 讨论性信息素成分在载体中的释放速率除了取决于其自身的挥发性外,另一个主要影响因素是释放载体。性信息素成分在诱芯载体中的氧化、异构化、释放比例和释放速率的变化都是影响仿生诱芯生物活性的重要因素。本试验不采用异构化速率较快的天然橡胶和释放速率较快的硅橡胶做性信息素的释放载体(Brown et al., 1986;荆慧荣等,1996;高伟等,2001),而是利用卤化丁基橡胶和天然橡胶构成的异构化速率较慢的复合橡胶载体(Witzgall et al., 1993)和聚乙烯管载体进行共轭二烯类性信息素成分释放速率和异构化方面的研究。另外,由于本试验是在风洞实验室中进行的,所以也不考虑聚乙烯管载体受光照影响而引起性信息素成分的氧化问题。本研究采用Porapak Q动态顶空吸附载体中的挥发性性信息素成分,进一步用气相色谱分析具有不同异构体和官能团性信息素成分释放速率的动态变化,该研究方法优于在不同时期收集性信息素载体,然后根据载体中信息素的残留量推测释放速率的方法。残留量法推测信息素在载体中的释放速率的缺点是不能确定该释放速率是由于性信息素成分的氧化、降解释放,还是其纯粹从载体上释放(Cork et al., 2008;Alfaro et al., 2009)。该方法也容易受萃取溶剂、萃取时间和温度等方面的影响。
根据化学原理,醛、醇和酯类性信息素成分的挥发性应该依次降低。但本试验中醇类性信息素成分的释放速率大于醛类,这说明信息素载体对不同性信息素成分的释放有很大影响。在袖口式复合橡胶载体中,点样后性信息素成分就以很高的释放速率挥发。而在聚乙烯管载体中,性信息素成分穿透聚乙烯管壁需要一定的时间,所以其挥发模式是先慢后快,到最后由于挥发和异构化使聚乙烯管中剩余的性信息素浓度不足以再释放到空气中时,造成了载体中性信息素成分的残留问题。例如,大谷蠹(Prostephanus truncates)聚集信息素成分是反-2-甲基-2-庚烯酸甲乙基酯和反, 反-2, 4-二甲基-2, 4-庚二烯酸甲乙基酯。Hodges等(2004)、Compton等(1997)对冰箱中存放了至少1周的聚乙烯管诱芯和刚配制的诱芯进行测量时发现,存放过的诱芯在释放开始的1-4小时内释放量由高到低发生了急剧变化,而刚配制的诱芯两天后才达到释放最大值,这说明冰箱中的性信息素成分已穿透诱芯管壁并达到释放平衡状态,暴露时平衡状态被打破而出现瞬间高释放的现象;刚配制的聚乙烯管诱芯需要2天的时间来穿透管壁,这与本研究结论相似。但是大谷蠹聚集信息素在聚乙烯管载体中释放速率过快的原因可能是由性信息素成分的差异引起的。
采用与本试验完全相同的聚乙烯管载体用称重法研究欧洲新松叶蜂(Neodiprion sertifer)性信息素成分(2S, 3S, 7S)-3, 7-二甲基-2-十五碳醇乙酸酯和欧洲赤松叶蜂(Diprion pini)性信息素成分(2S, 3R, 7R)-3, 7-二甲基-2-十三碳醇乙酸酯在其上的释放速率,得出2种松叶蜂性信息素成分在聚乙烯管载体上呈线性释放,符合线性回归方程y=a-bx(y:载体中的剩余量;a:初始点样量;b:释放速率;x:持效期),且释放速率的高低与温度和初始点样量有直接关系(Johansson et al., 2001)。例如,在平均加样量每聚乙烯管2.35 mg,18~21 ℃条件下,欧洲新松叶蜂性信息素成分能以10 μg·d-1的释放速率稳定释放100天;在27~30 ℃条件下,其能以15.6 μg·d-1的释放速率稳定释放45天。性信息素成分和研究方法方面的差异导致性信息素在聚乙烯管载体上的这种释放模式与本研究结论不同。另外,由于欧洲赤松叶蜂性信息素成分在橡胶载体上释放量过低而选用棉签(dental cotton roll)作释放载体(Bergstrm et al., 1995),但是棉签载体不具有长效的稳定释放速率,在干扰交配法防治松叶蜂时其最佳作用时间仅能持续30天(Anderbrant et al., 1995)。此结果与本试验中性信息素成分在聚乙烯管载体中的释放速率大而在橡胶载体上释放量低的结论相符。此外,该结论在其他人的研究中也得到了证实(Cork et al., 2001; Bergstrm et al., 1995)。例如,反11-十六碳烯醇在聚乙烯管载体和橡胶载体上的释放速率是反11-十六碳烯乙酸酯的2倍,这2种成分在聚乙烯管载体上的释放速率又是橡胶载体上的2倍(Cork et al., 2001)。
共轭二烯结构的性信息素成分异构化主要受载体和光照强度的影响(Brown et al., 1986)。在新配制的诱芯中共轭二烯成分就开始发生异构化,且在储存或应用的过程中异构化程度随着时间的延长而升高,这能显著影响仿生诱芯的生物活性。本研究结果显示:E5, Z7-12: OH在聚乙烯管载体和橡胶载体上都容易发生异构化,而Z5, E7-12: OH在复合橡胶载体上比聚乙烯管载体上更容易发生异构化。醛和酯类性信息素成分在聚乙烯管载体和复合橡胶载体上81天后的异构化率介于10%~25%,这部分解释了林间诱集松毛虫试验后期效果差的原因。聚乙烯管载体中性信息素成分的剩余量低于复合橡胶载体,此结果与2载体中信息素释放速率的快慢和持效期的长短结果相符。本试验中性信息素异构成分中E5, E7构型的量比Z5, Z7构型的量多,说明EE结构比ZZ结构更稳定、转换更容易,这与高伟等(2001)的研究结论一致。但Vrkoč等(1988) 研究结果显示:反7, 顺9-十二碳二烯乙酸酯(E7, Z9-12: OAc)在聚乙烯管载体上22天后异构化为E7, E9构型的程度仅为3%;在天然苯乙烯-丁二烯橡胶载体上(秋兰姆处理)29天后异构化为E7, E9构型的程度仅为1.3%。异构化效果明显低于本试验的结论,这与不同时间测定的异构化率有关。
松毛虫雌虫一般在进入暗期2 h后才开始有求偶行为,凌晨是性信息素释放的高峰期,且主要性信息素成分的释放量一般低于2 ng(Priesner et al.,1984;孔祥波等,2003)。本试验中6种性信息素成分81天后的释放速率仍远高于单头活雌蛾性信息素的释放量,理论上诱芯的持效期可达到3个多月。生产上松毛虫诱芯的持效期一般维持在40天左右,而落叶松毛虫(Dendrolimus superans)仿生诱芯的最佳持效期仅有20天,载体中性信息素成分释放比例的变化和发生异构化现象是降低仿生诱芯持效期和生物活性的重要因素。本文松毛虫性信息素成分在不同载体不同时间的释放速率和异构化效果的研究结论可为生产上种群监测或迷向法防控松毛虫的危害提供重要的理论指导。
| [] | 高伟, 赵成华, 施再喜. 2001. 马尾松毛虫性信息素在不同类型诱芯中的稳定性. 昆虫学报, 44(2): 213–220. |
| [] | 荆慧荣, 刘孟英. 1996. 几种橡胶载体对马尾松毛虫性信息素几何构型异构化的影响. 昆虫学报, 39(2): 126–131. |
| [] | 孔祥波, 赵成华, 孙永平, 等. 2003. 赤松毛虫性信息素次要组分的鉴定—组分、电生理和田间效果. 昆虫学报, 46(2): 131–137. DOI:10.7679/j.issn.2095-1353.2003.033 |
| [] | Alfaro C, Navarro-Llopis V, Primo J. 2009. Optimization of pheromone dispenser density for managing the rice striped stem borer, Chilo suppressalis (Walker), by mating disruption. Crop Protection, 28(7): 567–572. DOI:10.1016/j.cropro.2009.02.006 |
| [] | Anderbrant O, Löfqvist J, Hgberg H E, et al. 1995. Development of mating disruption for control of pine sawfly populations. Entomologia Experimentalis et Applicata, 74(1): 83–90. DOI:10.1111/eea.1995.74.issue-1 |
| [] | Bergstrm G, Wassgren A B, Anderbrant O, et al. 1995. Sex pheromone of the pine sawfly Diprion pini (Hymenoptera, Diprionidae): chemical identification, synthesis and biological activity. Cellutar and Molecular Life Sciences, 51(4): 370–380. DOI:10.1007/BF01928898 |
| [] | Brown D F, Mcdonough L M. 1986. Insect sex pheromones: formulations to increase the stability of conjugated dienes. Journal of Economic Entomology, 79: 922–927. DOI:10.1093/jee/79.4.922 |
| [] | Compton J A F, Floyd S, Acquaye K, et al. 1997. Comparison of three types of pheromone-baited flight traps for the Larger Grain Borer, Prostephanus truncatus (Horn). International Journal of Tropical Insect Science, 17(2): 205–211. DOI:10.1017/S1742758400016386 |
| [] | Cork A, Alam S N, Das A, et al. 2001. Female sex pheromone of brinjal fruit and shoot borer, Leucinodes orbonalis (Lepidoptera: Pyralidae): blend optimization. Journal of Chemical Ecology, 27(9): 1867–1877. DOI:10.1023/A:1010416927282 |
| [] | Cork A, De Souza K, Hall D R, et al. 2008. Development of PVC-resin-controlled release formulation for pheromones and use in mating disruption of yellow rice stem borer, Scirpophaga incertulas. Crop Protection, 27(2): 248–255. DOI:10.1016/j.cropro.2007.05.011 |
| [] | Hodges R J, Addo S, Farman D I, et al. 2004. Optimising pheromone lures and trapping methodology for Prostephanus truncatus (Horn)(Coleoptera: Bostrichidae). Journal of Stored Products Research, 40(4): 439–449. DOI:10.1016/S0022-474X(03)00047-X |
| [] | Ideses R, Shani A, Klug J T. 1982. Sex pheromone of European grapevine moth (Lobesia botrana) its chemical transformations in sunlight and heat. Journal of Chemical Ecology, 8(6): 973–980. DOI:10.1007/BF00987663 |
| [] | Ideses R, Shani A. 1988. Chemical protection of pheromones containing an internal conjugated diene system from isomerization and oxidation. Journal of Chemical Ecology, 14(8): 1657–1669. DOI:10.1007/BF01014550 |
| [] | Johansson B G, Anderbrant O, Simandl J, et al. 2001. Release rates for pine sawfly pheromones from two types of dispensers and phenology of Neodiprion sertifer. Journal of Chemical Ecology, 27(4): 733–745. DOI:10.1023/A:1010302002735 |
| [] | Kong X B, Zhao C H, Gao W. 2001. Identification of sex pheromones of four economically important species in genus Dendrolimus. Chinese Science Bulletin, 46(24): 2077–2081. DOI:10.1007/BF02901136 |
| [] | Kong X B, Zhang Z, Zhao C H, et al. 2007a. Female sex pheromone of the Yunnan pine caterpillar moth Dendrolimus houi: first (E, Z)-isomers in pheromone components of Dendrolimus spp. Journal of Chemical Ecology, 33(7): 1316–1327. DOI:10.1007/s10886-007-9313-2 |
| [] | Kong X B, Zhao C H, Wang R. 2007b. Sex pheromone of the larch caterpillar moth, Dendrolimus superans, from northeastern China. Entomologia Experimentalis et Applicata, 124(1): 37–44. DOI:10.1111/eea.2007.124.issue-1 |
| [] | Kong X B, Sun X L, Wang H B, et al. 2011. Identification of components of the female sex pheromone of the Simao pine caterpillar moth, Dendrolimus kikuchii Matsumura. Journal of Chemical Ecology, 37: 412–419. DOI:10.1007/s10886-011-9932-5 |
| [] | Millar J G. 2000. Polyene hydrocarbons and expoxides: a second major class of Lepidopteran sex attractant pheromones. Annual Review of Entomology, 45: 575–604. DOI:10.1146/annurev.ento.45.1.575 |
| [] | Priesner E, Bogenschültz H, Albert R, et al. 1984. Identification and field evaluation of a sex pheromone of the European pine moth.. Zeitschrift für Naturforschung: Section C, Biosciences, 39(11-12): 1192–1195. |
| [] | Rothschild G H L. 1979. A comparison of methods of dispensing synthetic sex pheromone for the control of oriental fruit moth, Cydia molesta (Busk)(Lepidoptera: Tortricidae), in Australia. Bulletin of Entomological Research, 69(1): 115–127. DOI:10.1017/S0007485300017958 |
| [] | Vrkoč J, Konečny K, Valterová I, et al. 1988. Rubber substrates and their influence on isomerization of conjugated dienes in pheromone dispensers. Journal of Chemical Ecology, 14(5): 1347–1358. DOI:10.1007/BF01020139 |
| [] | Weatherston L, Golub M A, Brooks T W, et al. 1981. Methodology for determining the release rates of pheromones from hollow fibers//Mitchell E R. Management of insect pests with semiochemcials, Concepts and Practice, Plenum Publishing, New York. |
| [] | Weatherston L, Miller D, Lavoie-Dornik J. 1985. Commercial hollow-fiber pheromone formulations: The degrading effect of sunlight on celcon fibers causing increased release rates of the active ingredient. Journal of Chemical Ecology, 11(12): 1631–1644. DOI:10.1007/BF01012117 |
| [] | Witzgall P, Bengtsson M, Unelius C R, et al. 1993. Attraction of pea moth Cydia nigricana F.(Lepidoptera: Tortricidae) to female sex pheromone (E, E)-8, 10-dodecadien-1-yl acetate, is inhibited by geometric isomers E, Z, Z, E, and Z, Z. Journal of Chemical Ecology, 19(9): 1917–1928. |