 2012, Vol. 48
2012, Vol. 48文章信息
- 薛瑶芹, 张文辉, 马莉薇, 马闯, 周建云
- Xuex Xue Yaoqin, Zhang Wenhui, Ma Liwei, Ma Chuang, Zhou Jianyun
- 不同生境下栓皮栎伐桩萌苗的生长特征及在种群更新中的作用
- Features of Quercus variabilis Stump Sprouting and Its Contribution to Population Regeneration in Different Habitats
- 林业科学, 2012, 48(7): 23-29.
- Scientia Silvae Sinicae, 2012, 48(7): 23-29.
-
文章历史
- 收稿日期:2011-07-08
- 修回日期:2011-12-07
-
作者相关文章



2. 西部环境与生态教育部重点实验室 杨凌 712100
2. Key Laboratory of Environment and Ecology in West China of Ministry of Education Yangling 712100
栓皮栎(Quercus variabilis)是我国暖温带落叶阔叶林和亚热带常绿落叶阔叶混交林的重要组成树种,分布于我国22个省区(张文辉等,2002)。栓皮栎经济和生态价值大,在分布区内主要作为提取丹宁、软木的资源树种,也是薪炭林和食用菌培育原料。多年的过度利用导致绝大多数天然栓皮栎林退化为残败次生林,生态功能和经济效益明显下降(张文辉等,2002)。栓皮栎有无性繁殖和有性生殖2种繁殖方式。栓皮栎种子含有大量的淀粉,在自然条件下,土壤中大部分的栓皮栎种子被动物搬运和取食,还有一部分霉变和未成熟,完好种子只占种子数量的16.49%~22.23%,林地中1年生幼苗数量与完好种子数量的比率为仅为0.11%~0.20%,而且实生苗在建成过程中死亡率较高(马莉薇等,2010),这些瓶颈的存在导致有性生殖在栓皮栎次生林尤其是干扰严重的次生林天然恢复中的作用有限。
无性繁殖是栓皮栎自然更新的重要途径,是有性生殖的重要补充(吴明作等,2001)。栓皮栎有强大的萌芽能力,树干基部、伐桩和根系上的休眠芽或不定芽均可以发育成萌生苗(卢志军,2002)。伐桩萌苗是栓皮栎无性繁殖的重要方式,而且相对于实生苗,伐桩萌苗的生长较为迅速(Lockhart et al., 2007),可以很快占据干扰所形成的林隙,在较短时间内恢复林地的郁闭度,恢复林地生态功能(Kammesheidt, 1998;Bond et al., 2001)。国内对于栓皮栎的无性繁殖尤其是伐桩萌苗的研究较少(吴明作等,2001;卢志军,2002),但对栎属其他植物的伐桩萌发特性进行了一定研究,指出萌生植株尤其是伐桩萌苗是辽东栎(Quercus wutaishanica)顺利通过更新瓶颈的一种手段,在种群更新中起一定作用(高贤明等,2001);立地因子,尤其是坡向、坡位等影响生境光照和温度的立地因子对辽东栎萌芽更新造成一定影响(伊力塔等,2006;2007)。
林窗常指由于森林中某一林冠乔木的死亡或者砍伐等原因造成林地上形成不连续的林中隙地(Watt, 1947)。林窗的形成导致了资源的再分配和微生境的异质性(Chazdon et al., 1991),使林窗与非林窗生境在光照条件、空气温度和湿度,甚至是微地形方面都有明显的差异,这些差异对林木幼苗定居和生长更新过程有重要影响(王彬等,2007)。林缘是森林与空地的结合部,其独特的环境梯度变化使森林表现出明显的边缘效应(刘金福,2003),这种效应对树木更新和生长产生重大影响。林内、林缘和林窗的生境异质性对栓皮栎伐桩萌苗更新的影响还未见报道。
针对不同生境中退化的栓皮栎种群而言,哪种繁殖方式是其主要的自然更新方式,能获得更好的效果,是栓皮栎种群恢复研究的关键所在。本文通过调查不同生境中(林内、林窗、林缘)栓皮栎幼苗的起源类型,对比栓皮栎不同起源幼苗的数量和重要值,阐明伐桩萌苗在不同生境的种群更新中的作用;通过对比不同生境下伐桩萌苗的生长状况,阐明环境条件对其生长的影响,为栓皮栎次生林改造提供理论依据和实践指导。
1 材料与方法 1.1 研究区选择及概况丁慧萍等(2007)采用分形理论和方法分析了黄龙山区、秦岭以北以及巴山北坡栓皮栎种群空间分布格局,发现在秦岭北坡海拔600~1 300 m内,栓皮栎种群占据生态空间的能力、聚集程度、种群密度等都是最大的。因此本研究以位于秦岭北坡的陕西省周至县楼观台地区(107°39′—108°37′E,33°42′—34°14′ N)为研究区域。
该区属温带大陆性季风气候,年均气温13.2 ℃,年均降水量650~800 mm,无霜期225天。土壤为山地褐土。栓皮栎为该地区乔木层的优势种,伴生种有20多种,主要包括:侧柏(Platycladus orientalis)、油松(Pinus tabulaeformis)、槲树(Quercus dentata)、槲栎(Quercus aliena)、黄栌(Cotinus coggygria)、绿叶胡枝子(Lespedeza buergeri)、美丽胡枝子(Lespedeza formosa)、榛(Corylus heterophylla)、黄蔷薇(Rosa hugonis)、小花扁担杆(Grewia biloba var. parviflora)、苔草(Carex tristachya)、麦冬(Ophiopogon japonicus)、紫菀(Aster tataricus)等。
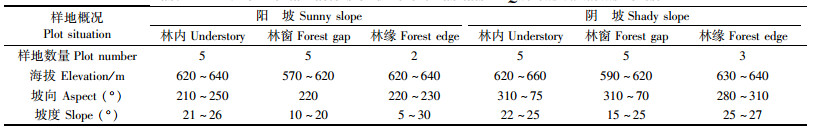
1.2 研究方法 1.2.1 样地设置本研究以该区域中分布在阴坡和阳坡林相较好的栓皮栎林为研究对象。该林分是1985年皆伐后形成的天然次生林,栓皮栎占林分组成90%以上,郁闭度为75%~80%。在2004年冬季有部分栓皮栎被砍伐,伐桩散布于整个林分。该林分距离居住区较近,有放牧痕迹。
2009年7—9月在不同小生境(林内、林缘、林窗)中布设固定样地。样地设置如下:林内样地10块(阳坡5块,阴坡5块),设置在林相整齐、林木分布均匀的地段,样地面积20 m×20 m(样地中心距林缘距离>40 m);林缘样地共5块(阳坡2块,阴坡3块),设置在森林边缘,即森林与农田或空地的交汇,面积为20 m×20 m(样地中心在林缘,由林缘向林内延伸);同时在林内寻找到10个林窗(阳坡5个,阴坡5个,样地中心距林缘距离>40 m)作为样地。在林下和林缘样地中延对角线设置3个5 m×5 m幼苗和灌木样方;林窗样地中共设置9个2 m×2 m幼苗和灌木样方,1个在林窗中央,其余8个分别沿林窗的长、短轴方向设置在林窗中央距边缘1/2处和林窗边缘处(宋新章等,2008)。草本样方设置同灌木样方,面积为1 m×1 m。
1.2.2 样地调查生境调查:用GPS(Magellan GPS 315)测定样地的海拔和地理坐标;用罗盘仪测定坡度和坡向,样地的立地因子概况见表 1;选择晴朗(前10天无雨)的天气,在3个生境的样地内,分别于10: 00,12: 00,14: 00,16: 00测定各样地中心地表以上0.5 m处的空气温度、湿度(DHM2型通风干湿温度计)、地表以上0.5 m处的光照强度(ZDS-10型照度计)。沿林内、林缘样地的对角线和林窗样地的长轴均匀布置3个土壤取样点,取地表以下0~15 cm混合土样,密封后带回室内,测定土壤含水量(马莉薇等,2010)。
|
|
草本、灌木调查:在灌木样方中调查每种灌木的数量、高度和盖度,在草本样方内调查草本植物,的数量、高度和温度,同时调查枯枝落叶厚度。
幼苗调查:在每一幼苗样方中调查所有高度≤2.5 m的栓皮栎幼苗,调查内容包括年龄、起源(实生、伐桩萌苗、干基萌苗、根系萌苗)、基径、高度、冠幅和叶片数,幼苗枯死则如实记录。样地中幼苗的年龄通过主茎上的芽鳞痕确定。伐桩上每一株萌苗均进行调查。
地上生物量和叶面积调查:在每一样地中选择直径相近的伐桩3~5个,在每个伐桩上取3~5株高度不同的萌苗作为标准株,将萌苗从其与桩基联结处取下,确定其年龄后,带回实验室。选取每株萌苗的各个方向上接近顶端的成熟叶片3~5枚,用EPSON PERFECTI ON 4870扫描仪将叶片扫描成图片文件,再用WinFOLIA 2004a软件分析叶面积(leaf area,LA)(祁建等,2008)。扫描叶片后,在80 ℃下将植株和取下来的叶片烘干至恒量,称量每株萌苗干物质质量,合并同一生境中同一年龄萌苗地上部分生物量数据,取其平均值。
1.2.3 数据计算及统计伐桩萌苗和干基萌苗均有在一个伐桩或母树上可生长多个个体的特征,如果仅仅以数量来论定其在种群更新中的作用是不恰当的,因此本研究采用重要值来判断不同起源幼苗在种群更新中的作用。
重要值计算公式为:(Kammesheidt,1998;宋永昌,2001)。
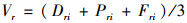
|
式中:Dri为相对密度,Pri为相对显著度,Fri为相对频度。
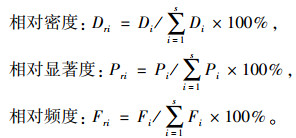
|
式中:Di为第i种起源幼苗的密度,Pi为第i种起源所有幼苗的基面积,Fi为第i种起源幼苗的频度。
使用SPSS 17.0对不同生境下栓皮栎幼苗的年龄组成、幼苗的生长指标、生境条件进行单因素方差分析(One-way ANOVA)和多重比较,在α=0.05水平上检验差异的显著性。应用Origin 8.0进行做图。
2 结果与分析 2.1 栓皮栎幼苗类型及其在种群更新中的地位通过对样地中幼苗的仔细调查发现,在该区域内,只有实生苗、伐桩萌苗和主干基部萌苗,根系萌苗并没有在该区域出现(表 2)。
|
|

通过对不同生境下不同类型幼苗的基面积对比可以发现:1)同一生境内,基面积均表现为伐桩萌苗>实生苗>干基萌苗;2)相同起源的幼苗,基面积表现为:林窗>林缘>林内,林缘和林内伐桩萌苗的基面积分别只有林窗的38.0%和24.9%。
2.1.2 重要值3个小生境中,重要值的变化趋势与基面积一致,均为伐桩萌苗>实生苗>干基萌苗,表明3个生境中均存在有性和无性2种更新策略,但伐桩萌苗是主要的更新方式(表 2)。不同小生境中,伐桩萌苗对种群更新的作用不同:在林内,伐桩萌苗和实生苗重要值的差异最小,2种更新方式占有同样的地位;在林缘,二者的重要值差异最大,实生苗的重要值仅为伐桩萌苗的51.6%;林窗中二者重要值差异介于中间,实生苗的重要值为伐桩萌苗的71.3%,伐桩萌苗占主要地位。
2.1.3 年龄结构通过比较各生境中不同起源幼苗的年龄结构(图 1)可以发现:1)林窗和林缘中,各年龄的伐桩萌苗数量均显著大于实生苗和干基萌苗(P < 0.05)(图 1B, 1C),林内1年生伐桩萌苗数量显著少于同龄实生苗(P < 0.05)(图 1A);2)实生苗的年龄结构在各生境中均为随着年龄增加,幼苗数量逐渐减少,不同生境中实生苗数量为林缘和林窗>林内,但1~2年生实生苗数量林缘>林窗,3~5年生则相反;3)伐桩萌苗的年龄结构为1~4年生萌苗数量较多且较稳定,5年生伐桩萌苗数量最少(林缘仅7株·100 m-2),林窗和林缘的伐桩萌苗数量均大于林内,只有5年生伐桩萌苗数量为林内>林缘;4)干基萌苗的年龄结构缺少1~2年生萌苗,萌苗数量为林缘>林内,林窗内没有干基萌苗。

|
图 1 不同生境中不同起源幼苗的年龄结构 Fig.1 Age structure of different original young individuals in different habitats 数据为平均值±标准误差。A为林内,B为林窗,C为林缘。不同字母表示数据在不同起源幼苗间差异显著。 Data are mean value ± standard error (SE). Different letters indicate significant difference among different origin young individuals. A means age structures in understory, B means age structures in forest gap, C means age structures in forest edge. |
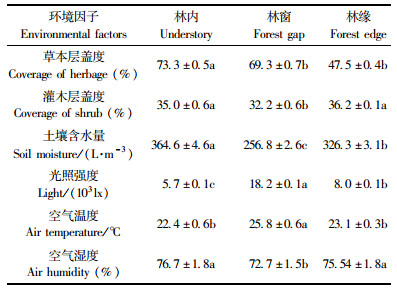
对林窗、林缘和林内的环境因子的单因素方差分析及多重比较表明:3种生境中的枯枝落叶层厚度、草本层盖度等环境因子间均存在显著性差异(P < 0.05)(表 3)。林窗的光照强度和空气温度显著大于非林窗生境(林内,林缘)(P < 0.05),但其空气湿度、土壤水分和灌木层盖度则相反(表 3),这充分说明林窗、林缘、林内生境存在较大的异质性。
|
|
每桩萌苗数量可以显示伐桩的萌发能力(Rydberg, 2000)。林窗和林缘的每桩萌苗数量显著高于林内(P < 0.05)(图 2),表明林窗和林缘有利于伐桩萌发。

|
图 2 不同生境下每桩萌苗数量对比 Fig.2 Numbers of sprouts per stump in different habitats |
林窗、林缘、林内的无机环境和生物环境都存在不同程度的差异(表 3),萌苗之间的竞争程度也表现出不同。萌苗的枯死率在林内、林窗及林缘间有显著差别(P < 0.05),林内萌苗的枯死率最低(12.5%),林窗萌苗的枯死率最高(20.3%),林缘介于中间(15.0%)(图 3)。虽然林窗萌苗数量较多[139株·(10 m)-2],但该生境内萌苗枯死数量也最多。

|
图 3 不同生境下萌苗的枯死率 Fig.3 Mortality rates of stump sprouts in different habitats |
在不同小生境内,萌苗基径、高度、叶面积以及地上生物量的生长趋势均与年龄呈正相关,年龄越大,平均基径、高度、叶面积和地上生物量也越大(图 4)。

|
图 4 不同生境中伐桩萌苗的生长情况 Fig.4 Growth features of Q. variabilis stump sprouts with different age in different habitats |
萌苗基径和萌苗地上部分生物量均表现出林窗>林缘>林下,这种差异也随着萌苗年龄的增长而变大,1~2年生幼苗基径在生境间差异不显著(P > 0.05),3~5年生萌苗基径林窗显著大于林下(P < 0.05)(图 4A,4D)。
萌苗高度和萌苗叶面积表现出一致的趋势,即林窗>林缘>林下,但萌苗高度和叶面积在生境间的差异随着年龄增长从显著(1~4年生,P < 0.05)变为不显著(5年生,P > 0.05),林窗中1年生萌苗的高度比林下高39.6%,叶面积大57.1%,而5年生萌苗的相应数值为21.2%和2.3%,1年生萌苗对微生境异质性的响应较为强烈。
以上结果表明:小生境的异质性导致各年龄萌苗的生长状况不同,但总体而言,萌苗在林窗中生长最好,其高度、基径、叶面积和地上生物量均大于林缘和林内萌苗。
3 讨论栓皮栎可以通过有性生殖和无性繁殖2种方式进行种群更新,通过有性生殖产生的实生苗分布到种子可以扩散到的地方,生境的多样性有利于实生苗适应环境的变化,而且实生苗对维持及扩展种群基因多样性和种群规模有重要意义(张文辉等,2002),但是实生苗定居过程中较高的死亡率以及较慢的生长速度(马莉薇等,2010)均影响其在种群更新尤其是受干扰种群恢复中的作用。萌生苗的分布受制于被伐母树,但其生长速度比实生幼苗快很多,例如,林窗中1年生、5年生实生苗高度为30 cm和100 cm(马莉薇等,2010),但同一生境中的伐桩萌苗高度分别为42 cm和128 cm(图 4B), 因此萌生苗在快速占据被伐母树的生态位以及覆盖干扰所形成的林隙或者空地方面优于实生苗(Bond et al., 2009)。实生苗和萌生苗对于种群的发展都是必不可少的,但二者在种群更新中的地位在不同生境中有不同表现。Bellingham等(2000)的模型指出在生产力低下的林地中,无性繁殖实际上优于有性生殖,而在生产力高的林地中则相反(Bellingham等(2000))。在干扰较大的环境中,栓皮栎属于潜在的r对策者,即种群依靠萌芽生殖产生巨大的幼苗库,以弥补种子繁殖能力有限、个体存活率低的不足,从而可以在较短时间内占据优势地位,排斥其他树种,形成较大密度的纯林(吴明作等,2001)。本研究中,林内栓皮栎的更新以伐桩萌苗和实生苗为主,而在林窗和林缘,栓皮栎种群更新主要依靠无性繁殖的伐桩萌苗,与Bellingham等(2000)和吴明作等(2001)的结论类似。林窗中实生幼苗在种群更新中的地位较低可能与动物干扰等原因造成的林窗种子库中栓皮栎完好种子数量少也有关系(马莉薇等,2011)。幼苗年龄结构与种群更新有密不可分的关系,在3个生境中,实生苗有完整的进展型幼苗库,表明实生苗能够持续发展(张文辉等,2003),而伐桩萌苗的年龄结构则是3~4年生萌苗的数量较多,1年生和5年生萌苗数量减少,说明伐桩的萌发能力和萌苗保存能力都在下降,其在种群更新中的地位能否继续保持需要进一步的跟踪研究。
一般来说,栎类植物的萌苗一般出现在植物体的3个地方,即伐桩、主干破损处和根系(Suh et al., 1998;Mostacedo et al., 2008)。卢志军(2002)也通过研究指出栓皮栎有3种萌苗类型,即伐桩萌苗,主干基部萌苗和根系萌苗。本研究区域中,栓皮栎只有2种萌苗,即伐桩萌苗和干基萌苗,而且伐桩萌苗的基面积和重要值远大于干基萌苗,而且各年龄的伐桩萌苗数量也多于干基萌苗,是该区域主要的无性繁殖方式。Mostacedo等(2008)对玻利维亚热带森林中31个树种的根系和伐桩萌苗进行的研究也指出,伐桩萌苗在森林的更新中占有主要地位。高贤明等(2002)也认为伐桩萌苗是辽东栎顺利通过更新瓶颈的有效手段。本研究中干基萌苗只出现在林缘和林内,而没有在林窗出现,可能是因为干基萌苗的出现和主干破损是紧密相关的,而林下和林缘植物遭受林冠层掉落物造成损伤的几率远远大于林窗(Lasso et al., 2009)。另外,伐桩萌苗的竞争激烈程度远远大于干基萌苗,到底有多少伐桩萌苗可以继续存活,尤其是5年以后的存活率以及进入主林层的几率还需要长时间的跟踪监测。
伐桩萌苗的建成和生长取决于许多因素,但光照条件是对其影响最大(Lockhart et al., 2007; 伊力塔等,2006)。Gardiner等(1997)、Lockhart等(2007)和Atwood等(2009)对不同光照条件下水栎(Quercus nigra)、樱皮栎(Quercus pagoda)、矮栎(Quercus prinus)和白栎(Quercus alba)的伐桩萌发特性研究证明光照充足的环境有利于伐桩萌苗的产生。Gardiner等(1997)对轻度和重度间伐下水栎伐桩萌苗的直径和高度生长研究指出:林冠层郁闭度越高,萌苗高度和直径越小,需要附加间伐以促进萌苗生长。林窗的形成可以改善环境的光照条件(张远彬等,2006),本研究中林窗的光照、温度和空气湿度显著高于林缘和林内,而林窗中伐桩的萌苗能力和萌苗生长均优于林内和林缘,与以上研究结论一致。林窗中伐桩萌苗地上部分生物量较大,表明光照强的环境有利于幼苗的生物量累积(马莉薇等,2010)。林窗萌苗枯死率(20.3%)高于林内(12.5%),这可能是由于林窗内光照条件强,温度高,土壤水分不足(表 3),再加上林窗和林缘每桩萌苗数量大于林内(图 2),萌苗间竞争激烈,导致萌苗死亡率较高。由于该林分靠近居民生活区,林缘的人为干扰大于林内和林窗,也造成一部分萌苗折断和死亡,因此林缘的萌苗枯死率(15%)也较高。
4 结论研究区域中,栓皮栎有3种更新幼苗:实生苗、伐桩萌苗和干基萌苗。在林内,种群更新以实生苗和伐桩萌苗为主;在林窗和林缘中,伐桩萌苗在种群更新中占主要地位。
小生境影响伐桩的萌苗能力、萌苗的保存率和生长。林窗和林缘伐桩萌苗能力较强,但枯死率也较高;林窗和林缘的伐桩萌苗生长优于林内。
不同年龄的萌生苗生长对生境异质性的响应不同,1~4年生萌苗的高生长和叶面积对环境条件响应强烈,3~5年生萌苗基径和地上生物量对环境异质性的响应较为强烈。
林窗的存在对栓皮栎种群的伐桩萌芽有利,在更新不良的栓皮栎次生林中可以适当间伐生长不良的栓皮栎个体,开辟林窗,有利于栓皮栎种群的伐桩萌芽更新。
| [] | 丁慧萍, 钱克红, 杨运经. 2007. 陕西栓皮栎种群分布格局的分形特征. 西北林学院学报, 22(6): 7–9. |
| [] | 高贤明, 王巍, 杜晓军. 2001. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 25(6): 673–678. |
| [] | 刘金福. 2003. 格式栲群落林窗边缘效应研究. 应用生态学报, 14(9): 1421–1426. |
| [] | 卢志军. 2002. 栓皮栎种群生殖生态学研究 . 杨凌, 西北农林科技大学出版社. |
| [] | 马莉薇, 张文辉, 薛瑶芹, 等. 2010. 秦岭北坡不同生境栓皮栎实生苗生长及其影响因素. 生态学报, 30(23): 6512–6520. |
| [] | 宋新章, 张智婷, 肖文发, 等. 2008. 长白山杨桦次生林采伐林隙幼苗更新动态. 林业科学, 44(3): 13–20. DOI:10.11707/j.1001-7488.20080307 |
| [] | 宋永昌. 2001. 植物生态学 . 上海, 华东师范大学出版社. |
| [] | 祁建, 马克明, 张育新. 2008. 北京东灵山不同坡位辽东栎叶属性的比较. 生态学报, 28(1): 122–128. |
| [] | 王彬, 王辉, 杨君珑, 等. 2007. 子午岭次生油松林主要乔木树种的更新特点. 浙江林学院学报, 24(5): 559–563. |
| [] | 吴明作, 刘玉萃, 姜志林. 2001. 栓皮栎种群生殖生态与稳定性机制研究. 生态学报, 21(2): 225–230. |
| [] | 伊力塔, 韩海荣, 马钦彦, 等. 2006. 灵空山辽东栎萌芽更新的灰色关联分析. 山西林业科技(1): 23–25. |
| [] | 伊力塔, 韩海荣. 2007. 山西灵空山林区辽东栎萌芽更新规律研究. 林业资源管理(4): 57–61. |
| [] | 张文辉, 卢志军. 2002. 栓皮栎种群的生物学生态学特性和地理分布研究. 西北植物学报, 22(5): 1093–1101. |
| [] | 张文辉, 卢志军, 李景侠, 等. 2003. 秦岭北坡栓皮栎种群动态的研究. 应用生态学报, 14(9): 1427–1432. |
| [] | 张远彬, 王开运, 鲜骏仁. 2006. 岷江冷杉林林窗小气候及其对不同龄级岷江冷杉幼苗生长的影响. 植物生态学报, 30(9): 942–946. |
| [] | Atwood C J, Fox T R, Loftis D L. 2009. Effects of alternative silviculture on stump sprouting in the southern Appalachians. Forest Ecology and Management, 257(4): 1305–1313. DOI:10.1016/j.foreco.2008.11.028 |
| [] | Bellingham P J, Sparrow A D. 2000. Resprouting as a life history strategy in woody plant communities. Oikos, 89(2): 409–416. DOI:10.1034/j.1600-0706.2000.890224.x |
| [] | Bond W J, Midgley J J. 2001. Ecology of sprouting in woody plants: the persistence niche. Trend in Ecology & Evolution, 16(1): 45–51. |
| [] | Chazdon R L, Pearcy R W. 1991. The importance of sunflecks for forest understory plants. Bio Sci, 41(11): 760–766. |
| [] | Gardiner E S, Helmig L. 1997. Development of water oak stump sprouts under a partial overstory. New Forest, 14(1): 55–62. DOI:10.1023/A:1006502107495 |
| [] | Kammesheidt L. 1998. The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-and-burn agriculture in Eastern Paraguay. Plant Ecology, 139(2): 155–165. DOI:10.1023/A:1009763402998 |
| [] | Lasso E, Engelbrecht B M J, Dalling J W. 2009. When sex is not enough: ecological correlates of resprouting capacity in congeneric tropical forest shrubs. Oecologia, 161(1): 43–56. DOI:10.1007/s00442-009-1353-6 |
| [] | Lockhart B R, Chambers J L. 2007. Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley, USA. New Forests, 33(2): 183–192. DOI:10.1007/s11056-006-9022-7 |
| [] | Mostacedo B, Putz F E, Fredericksen T S, et al. 2009. Contributions of root and stump sprouts to natural regeneration of a logged tropical dry forest in Bolivia. Forest Ecology and Management, 258(6): 978–985. DOI:10.1016/j.foreco.2008.09.059 |
| [] | Rydberg D. 2000. Initial sprouting, growth and mortality of European aspen and birch after selective coppicing in central Sweden. Forest Ecology and Management, 130(1/3): 27–35. |
| [] | Suh M H, Lee D K. 1998. Stand structure and regeneration of Quercus mongolica forests in Korea. Forest Ecology and Management, 106(1): 27–34. DOI:10.1016/S0378-1127(97)00236-3 |
| [] | Watt A S. 1947. Pattern and process in the plant communities. Journal of Ecology, 35(1/2): 1–22. DOI:10.2307/2256497 |