 2012, Vol. 48
2012, Vol. 48文章信息
- 容丽, 王世杰, 俞国松, 邓晓琪, 冉景丞
- Rong Li, Wang Shijie, Yu Guosong, Deng Xiaoqi, Ran Jingcheng
- 荔波喀斯特森林4种木本植物水分来源的稳定同位素分析
- Stable Isotope Analysis of Water Sources of Four Woody Species in the Libo Karst Forest
- 林业科学, 2012, 48(7): 14-22.
- Scientia Silvae Sinicae, 2012, 48(7): 14-22.
-
文章历史
- 收稿日期:2011-05-23
- 修回日期:2011-11-10
-
作者相关文章



2. 中国科学院地球化学研究所环境地球化学国家重点实验室 贵阳 550002;
3. 河海大学水文水资源与水利工程科学国家重点实验室 南京 210098;
4. 贵州茂兰国家级自然保护区管理局 荔波 558400
2. State Key Laboratory of Environmental Geochemistry, Institute of Geochemistry, Chinese Academy of Sciences Guiyang 550002;
3. State Key Laboratory of Hydrology-Water Resources and Hydraulic Engineering, Hohai University Nanjing 210098;
4. Management Bureau of Guizhou Maolan Nature Reserve Libo 558400
中国南方喀斯特高原区虽然地处亚热带季风性湿润气候区,但由于喀斯特充分发育,土层浅薄,土被不连续,地表保水能力差,大气降水入渗系数大,漏失严重,加之降水时空分布不均,使水分成为喀斯特山区植物生长的主要限制因子。喀斯特植物水分的确切来源一直是困扰植物学界的一个问题,运用植物根系吸收水分时不发生同位素分馏这一原理,采用木质部水分同位素分析有助于了解植物利用的水源,探讨生态系统中植物对水分的竞争和利用策略,可以更好地了解生态系统的结构与功能。
对陆生植物而言,传统方法很难确定植物吸收哪层土壤水分,尽管根系是植物的吸水器官,但植物根系的存在并不意味着这些根在水分吸收方面表现活跃(Flanagan et al., 1992;Thorburn et al., 1994)。同时,喀斯特地区建立根系模型需要非常巨大的人力成本,使得该区域仅有极少量关于根系分布的经验数据(Schenk et al., 2005)。喀斯特浅薄的土层和坚硬的基岩及石灰岩所特有的呈网状的裂隙、石缝、石沟使得该区根系分布模型可能与其他常态地貌下的根系模型有较大差异。在热带地区,根系长度与降水量呈负相关,与干旱时间呈正相关(Schenk et al., 2002),根系分布对木本植物功能的维持起着关键作用,喀斯特树种在没有灌溉的条件下能支持物种度过严酷的干旱时期,显示喀斯特植物可能通过根系的伸展获取稳定的水源(Benjamin et al., 2001)。
直接测定地下的生态过程是十分困难的(Ehleringer et al., 1992)。运用稳定同位素技术,通过测定植物木质部水分和不同土层水分的D和18O,可以帮助研究者判断植物根系有效吸水区域(Ehleringer et al., 1991; Brunel et al., 1995; Thorburn et al., 1995; Mensforth et al., 1996)。目前,氢氧同位素技术在森林植被(Ish-Shalom et al., 1992; Ewe et al., 1999; Atsuko et al., 2002;Pefluelas et al., 2003)、河岸植被(Dawson et al., 1991; Smith et al., 1991; Busch et al., 1992; Thorburn et al., 1993; Mensforth et al., 1994; Dawson et al., 1996; Jolly et al., 1996)、海岸植被(Valentini et al., 1995; Slavich et al., 1999)、沙漠植被及农田植被(Smith et al., 1997; Sekiya et al., 2002),以及具风化岩层的喀斯特植被(Querejeta et al., 2006;2007; McCole et al., 2007)中都得到了不同程度的应用。
研究区成土母岩为纯质碳酸盐岩,且形成年代老,结构致密,空隙度很低( < 3%),酸不溶物含量很低( < 4%)(中国科学院地质研究所岩溶研究组, 1987; 袁道先等,1994),几乎不存在疏松的风化岩层,表层根系生态空间狭小,使得土壤层以下广厚的岩层成为植被根系生长巨大的生态空间。在喀斯特的岩石层面裂隙发育,形成了与地表关系密切的地表岩溶带,喀斯特表层岩溶带水正是位于渗流带上部,土层以下30 m左右深度内基岩风化裂隙中作侧向运动的水流,其构成一个上层悬挂的饱和带(Williams,1983)。由于水分在表层岩溶带运行缓慢,运行速度在10-7~10-4m·d-1(Williams,1985),形成了稳定含水层,该层称为喀斯特表层岩溶带水(subcutaneous flow)。
假设当浅薄土层的水分不能支撑植物水分需要的时候,喀斯特表层岩溶带水成为植物的稳定水源。因此,设定本研究目标为:1)测定植物在利用土壤水的同时是否利用及利用了多少比例的表层岩溶带的喀斯特表层岩溶带水;2)测定林地植物清晨水势,并探讨植物利用水源与水势的关系;3)探讨不同功能群植物利用水源是否存在差别。
1 研究区概况 1.1 区域概况研究区位于贵州茂兰国家级自然保护区缓冲区,地处云贵高原南缘,是云贵高原向广西丘陵盆地的过渡地带,地理位置为107°52′10″—108°05′40″ E,25°09′20″—25°20′50″ N, 海拔850~1 100 m。该区是目前世界上同纬度地区残存下来仅有的、原生性强、相对稳定的岩溶森林生态系统,也是岩溶区原生性森林分布面积最大的地区(王世杰等,2007),区内多数地段是中亚热带原生性喀斯特森林,为常绿落叶阔叶混交林,也有不同退化程度的演替群落,对退化群落自然恢复的研究有很强的代表性(刘攀峰等,2008)。基岩为纯质石灰岩与白云岩。土壤不成片,呈零星分布,基岩出露面积高达80%。植被多生长于石缝、石沟、小土坑、微台地上。地貌类型为峰丛洼地,森林及灌丛植被多分布于坡面上,坡脚具很多泉点。本区属于中亚热带季风湿润气候地区,年均温15.3 ℃,1月平均气温为5.2 ℃,7月平均气温23.5 ℃,生长期237天,全年降水量1 752.5 mm,集中分布于5—10月,夏季半年(5—9月)的降水量多达1 420 mm,占全年总降水量的81%,年平均相对湿度83%。研究区降水时空分布极为不均,水源主要依靠大气降水的补给。

|
图 1 茂兰喀斯特森林研究区1995—2008年气候状况 Fig.1 Climatic conditions in study araea in Maolan during 1995—2008 |
样地土被分布不连续,地表石灰质基岩裸露,岩石表面零星分布有极薄的土壤和枯枝落叶,大量风化残余物存在于岩石构造裂隙中, 植物根系可以在这些裂隙中生长, 地上部分形成连续的植被层。选取的喀斯特原生乔木林是该地带的顶级森林群落,喀斯特次生林是演替中间阶段的森林群落,而灌木林是演替初级阶段群落,每群落类型选择9~11个样方。群落植被概况见表 1,植被树种主要是一些适应性强的乡土种,以耐旱、喜钙类型为主。
|
|
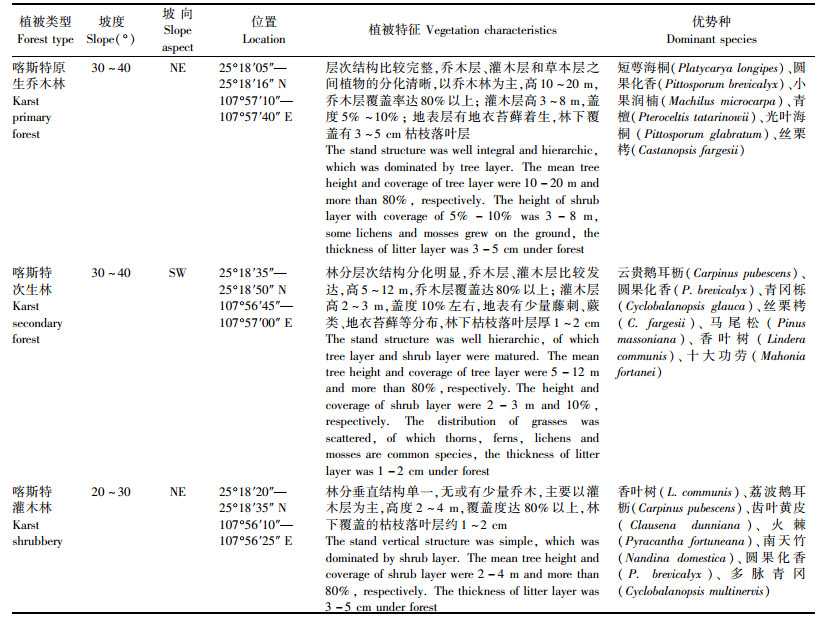
物种采样:在设置的3个样地,每样地设置的9~11个30 m×30 m样方中,选取香叶树(Lindera communis)、火棘(Pyracantha fortuneana)、齿叶黄皮(Clausena dunniana) 3种常绿灌木,另外选择1种落叶乔木幼树圆果化香(Platycarya longipes)作为对照。为便于比较,样地植株均选择胸径/基径(乔木约10 cm,灌木约4 cm)大小基本一致的个体,每种植物选择3~5株,同时记录样株生境。取样时间为少雨季节(2007年10月)和雨季中期和末期(2007年8月)。植物开始蒸腾作用的上午中间时段,取完全栓化没有叶片小枝,直径在0.5~1.0 cm,长8~15 cm,距离枝端几厘米的茎,避免其木质部水分受到叶片水分的干扰。植物样立即放入定制玻璃管中,并立即用带有O型密封圈的螺纹盖盖紧,防止蒸发(Bollard, 1960)。
土样的采集:在植物样冠幅内随机采集土壤混合样,从地表 0~10,10~30,30~50,>50 cm分别取土样(约30 g)混合完全后立即放入定制螺纹玻璃瓶中密封以防蒸发。样品立即冷冻保存。
水样的采集:收集了研究区4个喀斯特表层泉以代表上层滞水带及该区浅层地下水的稳定同位素组成。同时在该区采样点附近设置3个固定点收集大气降水,雨水样及泉水样均取样后立即封装于10 mL左右的事先准备好的干净聚乙烯瓶中,表层增加少量矿物油,并用parafilm密封瓶口,冷藏(4 ℃)备用(Peñuelas et al., 2003)。
2.2 清晨水势测定植物清晨水势(PDWP)反映黎明前植物水分与土壤水分达到平衡的状态(Larcher, 1997)。在采样日测定植物清晨水势(3:00—6:00) (Snyder et al., 2000)。水势测定采用PSΨPRO八通道水势仪(Wescor INC.,US)。
2.3 植物和土壤水分的萃取及氢氧同位素的测定植物茎及土壤水的提取采用较常见的低温真空蒸馏法(Ehleringer et al., 1989, Dawson et al., 1991)。氧稳定同位素分析采用CO2-H2O平衡法(Epstein et al., 1953;Socki et al., 1992);氢同位素分析采用H2-H2O平衡法。萃取的土壤、植物木质部水样、表层岩溶带水及雨水水样采用环境地球化学国家重点实验室连续流质谱(continuous flow MS)Isoprime-GC进行测定,实验标准采用SMOW(标准平均海洋水),测量误差,δD小于4‰,δ18O小于0.15‰,δD和δ18O值由国际通用标准形式给出。
δD(‰)=[((D/H)sample-(D/H) standard)/(D/H)standard]×1 000,
δ18O(‰)=[((18O/16O)sample-(18O/16O)standard)/(18O/16O)standard]×1 000。
2.4 水分来源比例的测算Walker等(1991)提出如何通过测定土壤水及植物的同位素值建立植物根系的水分吸收机制的假说。Brunel等(1995)应用这个概念发展了通过测定土壤水和植物木质部的同位素值,确定2层土壤水贡献的模型。这一模型能扩展到通过测定δD和δ18O的值确定3种水源,根据研究地的特点,确定植物3种可能的水源,如本研究中的土壤表层水、土壤下层水、喀斯特表层岩溶带水。具体模型参考Brunel等(1995)。
3 结果与讨论 3.1 植物清晨水势植物清晨水势测定结果表明:无论雨季或旱季,各树种间水势差异不显著(雨季:P=0.424;旱季:P=0.076),但旱季物种间水势差异大于雨季。香叶树的清晨水势最低,吸水能力最强,与何纪星等(1997)的研究结果一致。就旱季-雨季水势变化而言,乔木树种和灌木树种旱季和雨季的水势差没有显著差异,总体呈现相对稳定的状态,乔木种(圆果化香)清晨水势变化幅度略大于所测灌木种,且圆果化香的清晨水势最低,一般认为这是根系分布较浅、没有稳定水源所造成的(Schwinning, 2008)。
|
|

研究区地处中国南部季风性湿润气候区,降水的δD与δ18O同位素的雨量效应和季节效应明显,夏半年(通常5—10月) δD与δ18O值偏负, 冬半年(11月—次年4月) δD与δ18O值偏正(Liu et al., 1997; Tu et al., 2004)。受季节性降水(Gat et al., 1981)和蒸发影响,不同取样时间测定样品δ18O和δD值总体差异显著(P < 0.001)(图 2)。表层岩溶带水、土壤水及植物小枝水分斜率低于当地降水线,表明当地水分来源于雨水,同时降水在转变为各类水的过程中,蒸发作用导致了一定分馏。

|
图 2 各水δD及δ18O值与当地大气降水线LMWL的关系 Fig.2 Relationships between δD, δ18O values of different water and LMWL |
其中,表层岩溶带水位于渗流带上部,继承了一般地下水的所有性质(Willliams, 1985),由于降水对表层岩溶带水迅速的补给作用,渗流带上部的水源继承了雨水的同位素特性(图 2),即存在明显的季节差异和雨量效应。
不同土层梯度中土壤水分的δ18O的变化受近期降水与土壤原有水分同位素差异的联合影响(Ehleringer et al., 1992)。土壤水分的蒸发导致了土表水D,18O的富集(Allison et al., 1983),表层土壤水分的同位素组成往往比下层偏正(Smith et al., 1997; Walker et al., 1991; Sterberg et al., 1991)。喀斯特地区土层浅薄,土温变化强烈(容丽等, 2006),表层(0~10 cm)受大气影响显著,D,18O的富集;下层(>10 cm),土温变化趋缓,D,18O偏负于表层,各层内同位素变化较小(罗维均,2007)。
不同水源与植物小枝的δD及δ18O同位素相关性分析显示:喀斯特植物木质部水、土壤水及表层岩溶带水的衰减趋势(斜率)相似,显示土壤水和表层岩溶带水均是植物可靠水源(图 3)。

|
图 3 样地不同水源与植物小枝的δD及δ18O的同位素特征 Fig.3 Characters of δD and δ18O values in twig xylem water and different water source in the study sites a.次生林样地Secondary forest; b.灌木林样地Shrubwoods; c.原生林样地Original forest. |
样地最大土深在40~60 cm,根据喀斯特地区土壤水同位素组成的差异特点,土层主要分2层:上层(0~10 cm),受降水蒸发影响,变化强烈。下层(>10 cm)土壤水分既受到大雨的影响,也在某种程度上受到地下水的影响,通常该层水分代表贮存雨水的性质(罗维均, 2007; Cramer et al., 1999)。经计算,不同水源对所测植物的水分贡献比例如图 4。

|
图 4 3个群落及4种木本植物对表土层(0~10 cm)、下土层(>10 cm)及表层岩溶带水提升及利用的比例 Fig.4 Calculated water uptake from two soil layers and the subcutaneous flow for the 4 species in secondary forest, shrubbery and virgin forest plots F1-1.湿季次生林样地Wet secondary forest; F1-2.干季次生林样地Dry secondary forest; F2-1.湿季灌木林样地Wet shrubwoods; F2-2.干季灌木林样地Dry shrubwoods; F3-1.湿季原生林样地Wet original forest; F3-2.干季原生林样地Dry original forest. S1-1.湿季香叶树样地Wet L. communis; S1-2.干季香叶树样地Dry L. communis; S2-1.湿季齿叶黄皮样地Wet C. dunniana; S2-2.干季齿叶黄皮样地Dry C. dunniana);S3-1.湿季火棘样地Wet P. fortuneana; S3-2.干季火棘样地Dry P.fortuneana; S4-1.湿季圆果化香样地Wet P. longipes; S4-2.干季圆果化香样地Dry P. longipes. |
通过模型计算(图 4a),总体而言,各群落类型中,土壤水和稳定含水的喀斯特表层岩溶带水都是植物主要水源。其中土壤水分(包括土表和土壤下层水)是植物水分的主要水源,且土壤下层水的贡献比例最大。植物生长旺季的8月,土壤下层水对各样地的贡献最高,其中次生林样地土表、土壤下层及表层岩溶带水的利用较均衡,比例分别为32%,34.35%和33.65%;相比较而言,灌木林和原生林样地植物土壤下层水利用比例更高(>50%);降水减少的10月,灌木林转而主要利用土壤表层水(40.32%),可能与灌木林内既有原大树被砍伐后根系较深的林木,也有新萌发的幼树,而幼树根系分布较浅,且进入干季,无效降水的频次( < 5 mm)增加,较开阔样地的表层土壤更容易获得滋润,土表水有时更易利用;而次生林和原生林两个乔木为主群落仍然主要利用土壤下层水(分别为62.89%和45.52%),但总体而言,土壤下层水的贡献比例均大于1/3,可认为无论是雨季的8月或降水减少的10月,土壤水分,特别是土壤下层水是荔波喀斯特植物的主要水源。
就表层岩溶带水的利用而言,8月灌木林和原生林表层岩溶带水用量明显小于次生林样地(灌木林为12.39%;原生林为9.71%),均为10%左右。这可能一方面由于次生林样地上游来水量较大(1号样地的泉水量大于其他2个样地),地下水位较浅,植物更容易利用;另一方面,可能与3个样地土地利用的历史有关,次生林样地为退耕后恢复的山地,而原生林样地受人类干扰较少,灌木林样地为砍伐所致,植物及土壤未被完全破坏,特别是地下根系部分保留了较完整的原生特性。降水减少的10月,次生林表现出与灌木林和原生林不完全一致的水源特征,灌木林和原生林利用了更多的表层岩溶带水(分别为24.26%和13.14%),而次生林为土壤下层水的利用比例增加(62.89%),这也暗合了次生林根系次生性较强,根系分布较浅的特征;同时,灌木林和原生林样地因未被彻底破坏,储存表层岩溶带水的裂隙长期被枯落物填充,裂隙中的表层岩溶带水消散较慢,而次生林由于曾被开垦为耕地,裂隙渗漏性较强,表层岩溶带水在干旱季节更容易因渗漏而下降较快,而使得表层岩溶带水对各植被类型样地的供应存在差异。
不同物种对水分利用呈现不同特点(图 4b),降水少的10月,香叶树表层岩溶带水利用比例明显提升至26.6%;如前文所述喀斯特表层岩溶带水受降水影响显著,少雨的10月,表层岩溶带水位下降,而香叶树仍然能充分地利用表层岩溶带水,表明香叶树根系相对较深,可下扎至较深的岩隙中获取充足水分;而相关研究也表明:香叶树叶片形态解剖特征的变异系数最小(何纪星等,1997a),这与香叶树能持续获得稳定的水源支持、免受环境变化的干扰有关。结合香叶树所具有的低细胞渗透势和强的细胞吸水能力(何纪星等,1997a),为保证植物的稳定生理状态提供了可能。
齿叶黄皮雨季的8月和10月土表水的用量分别为53.90%和47.35%,变化不大;而10月土壤下层水用量明显提升(从8月的29.9%增加到10月的42.2%),而表层岩溶带水用量在2个取样季节用量均不足1/5,且表层岩溶带水的贡献比例从8月的16.18%下降至10月的10.43%。显然,整个生长季,土壤水均是齿叶黄皮的主要水源,且随着降雨量减少,含水量相对稳定的土壤下层成为植物相对稳定的水源;齿叶黄皮根系下扎岩隙的程度不如香叶树,随着10月表层岩溶带水位的下降,利用表层岩溶带水的比例下降。
火棘无论在8月或10月均较稳定地以利用土壤下层水为主,用量大于50%;土表水贡献率分别为28.5%和31.55%,地下水在10月的利用也从8月的17.81%下降至10.14%;火棘水分利用方式与齿叶黄皮类似,以土壤水为主要水源,根系下扎岩隙的程度也不如香叶树,随着10月表层岩溶带水位的下降,利用表层岩溶带水的比例下降。
作为唯一乔木落叶树种的圆果化香对各层水分用量均比较均匀,但10月土表水用量从8月的24.73%增加至50.51%;而土壤下层水用量减小,从51.22%减至32.98%;表层岩溶带水的用量也从8月的24.05%下降至10月的16.51%。显然,作为可生长为高大落叶乔木的圆果化香而言,其根系不属深根型植物,主要利用的仍然是土壤水,且随着表层喀斯特水位的下降水源利用的比例降低。从其水分参数而言,其束缚水/自由水的比例是该区主要物种中最低的(何纪星等,1997b),即叶片的耐旱能力并不强。但圆果化香却以最有效的落叶这种休眠方式度过恶劣环境,10月,落叶物种已开始为进入休眠期做准备,叶片开始发黄,生理功能下降,蒸腾作用降低,物种对水分的利用减小;故植物表征为表层水分利用比例提高,可能与植物生理功能下降、吸水量下降有关,同时,圆果化香主要分布于土层比较浅薄的土面或直接生长于石面之上,也可能暗示圆果化香主要通过扩展根幅获取充分的生态空间,即具有广而浅的根系,更倾向于获得不定降水补给的表层土壤水分。
在所测4个物种中,火棘水源利用最为保守,无论雨季或干季均主要利用相对稳定的土壤下层水;而落叶的圆果化香在降水较少的10月,土表水利用比例更高,体现出这2种植物水源利用的灵活性,进入干季,无效降水的频次( < 5 mm)增加,表层土壤更容易获得滋润,Schwinning等(2005)认为,若不定的降水是比稳定的深层土壤水更为重要的环境水资源,则最理想的植物表现型将倾向于能够最大限度利用不定降水。反之,若深层土壤水更为重要,则植物表现型将倾向于能够最大限度利用深层土壤水(Ehleringer et al., 1991;Valentini et al., 1992;Donovan et al., 1994;Jackson et al., 1995)。
4 结论1) 研究结果显示:降水、土壤水及表层岩溶带水均属同一变化趋势,表明地处在亚热带季风气候区的研究区,降水丰沛,土壤水及表层岩溶带水均来源于雨水。
2) 土壤水是各植被类型的主要水源,但随旱季来临,灌木林和原生林对表层岩溶带水用量的有效比例提升。
3) 物种水分利用策略存在差异。在常绿种中,香叶树根系较深,对表层岩溶带水的依赖性较强;而齿叶黄皮和火棘主要利用土壤下层水;落叶种随旱季来临减少了水分利用,乔木落叶种圆果化香对表层岩溶带水用量减少,表层水的利用比例增高,结合圆果化香的生境分布特征及清晨较低水势的特点,暗示了圆果化香较浅的根系分布,水分提升能力小,难以利用随降水减少而水位下降的表层岩溶带水。
喀斯特木本植物对地表水源的利用是非常充分和灵活的,无论是表层、深层土壤水或具地下水性质的表层岩溶带水都为植物生长做出了贡献。森林和灌丛植物对于土壤水分条件变化的适应是不同的,研究喀斯特峡谷区森林植被中几种优势植物水分来源,某种程度上有利于解释和调控地表水文过程,改善喀斯特植物不同时期水分管理方式,有助于理解喀斯特地区植物的水分利用策略。
| [] | 何纪星, 姚立. 1997a. 茂兰喀斯特森林树种叶形态解剖特征研究//朱守谦. 喀斯特森林生态研究Ⅱ. 贵阳: 贵州科技出版社, 33-41. |
| [] | 何纪星, 朱守谦, 韦小丽. 1997b. 喀斯特森林树种的PV曲线研究//朱守谦. 喀斯特森林生态研究Ⅱ. 贵阳: 贵州科技出版社, 25-32. |
| [] | 罗维均. 2007. 贵州喀斯特洞穴的稳定同位素地球化学特征. 中国科学院地理化学研究所博士学位论文. |
| [] | 容丽, 王世杰, 杜雪莲. 2006. 喀斯特低热河谷石漠化区环境梯度的小气候效应:以贵州花江峡谷区小流域为例. 生态学杂志, 25(9): 1038–1043. |
| [] | 袁道先. 1994. 中国岩溶学. 北京, 地质出版社. |
| [] | 中国科学院地质研究所岩溶研究组. 1987. 中国岩溶研究. 北京: 科学出版社. |
| [] | Allison G B, Barnes C J, Hughes M W. 1983. The distribution of deuterium and 18O in dry soils. Experimental J Hydrol, 64(3): 377–397. |
| [] | Atsuko S, Nao Y, Daisuke N, et al. 2002. Importance of ermafrost as a source of water for plants in east Siberian taiga. Ecological Research, 17(2): 493–503. |
| [] | Bollard E G. 1960. Transport in the xylem. Ann Rev Plant Physiol, 11(1): 141–166. DOI:10.1146/annurev.pp.11.060160.001041 |
| [] | Benjamin T J, Montañez P I, Jimênez J J M, et al. 2001. Carbon, water and nutrient Xux in Maya homegardens in the Yucatán peninsula of México. Agroforet System, 53(1): 103–111. |
| [] | Busch D E, Ingraham N L, Smith S D. 1992. Water uptake in woody riparian phratophytes of the southwestern United States: a stable isotope study. Ecological Applications, 2(4): 450–459. DOI:10.2307/1941880 |
| [] | Brunel J P, Walker G R, Kennett-Smith A K. 1995. Field validation of isotopic procedures for determining source water used by plants in a semi-arid environment. J Hydrol, 167(3): 351–368. |
| [] | Cramer V A, Thorburn P J, Fraser G W. 1999. Transpiration and groundwater uptake from farm forest plots of Casuarina glauca and Eucalyptus camaldulensis in saline areas of southeast Queensland, Australia. Agricultural Water Management, 39(2): 187–204. |
| [] | Dawson T E, Ehleringe J R. 1991. Streamside trees that do not use stream water. Nature, 350(3): 335–337. |
| [] | Dawson T E, Pate J S. 1996. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation. Oecologia, 107(1): 13–20. DOI:10.1007/BF00582230 |
| [] | Dawson T E.Ehleringer J R.1998: Plants, isotopes and water use:a catchment scale perspective// Kendall C, McDonnell J J.Isotpe tracers in catchment hydrology.New York:Elsevier, 165-202. |
| [] | Ehleringer J R, Osmond C B. 1989. Stable isotopes//Pearcy R W, Ehleringer J, Mooney H A, et al. Plant physiological ecology: Field methods and instrumentation. London: Chapman & Hall, 281-300. |
| [] | Ehleringer J R, Phillips S L, Schuster W S F, et al. 1991. Differential utilization of summer rains by desert plants, implications for competition and climate change. Oecologia, 88(5): 430–434. |
| [] | Ehleringer J R, Dawson T E. 1992. Water uptake by plants:perspectives from stable isotope composition. Plant Cell and Environmen, 15(6): 1073–1082. |
| [] | Ehleringer J R. 1993. Carbon and water relations in desert plants: an isotopic perspective//Ehleringer J R, Hall A E, Farquhar G D, et al. Stable isotopes and plant carbonwater relations. San Diego: Academic Press: 155–172. |
| [] | Epstein S, Mayeda T. 1953. Variations of O-18 content of water from natural sources. Geochimica et Cosmochimica Acta, 42(2): 213–224. |
| [] | Ewe S M L, Sternberg L S L, Busch D E. 1999. Water-use patterns of woody species in pineland and hammock communities of South Florida. Forest Ecology and Management, 118(1): 139–148. |
| [] | Flanagan L B, Ehleringer J R, Marshall J D. 1992. Differential uptake of summer precipitation among co-occurring trees and shrubs in a pinyon-juniper woodland. Plant Cell Environ, 15(5): 831–836. |
| [] | Gat J R, Confiantini R. 1981. Stable isotope Hydrology Deuterium and Oxygen-18 in the Water Cycle. Vienna: International Atomic Energy Agency: 6–73. |
| [] | Ish-Shalom N, Sternberg L da S L, Ross M S, et al. 1992. Water utilization of tropical hardwood hammocks of the lower Florida Keys. Oecologia, 92(1): 108–112. DOI:10.1007/BF00317270 |
| [] | Jackson P C, Caveliser J, Goldstein G, et al. 1995. Partitioning of water resources among plants of a lowland tropical forest. Oecologia, 101(2): 197–203. DOI:10.1007/BF00317284 |
| [] | Jolly I D, Walker G R. 1996. Is the field water use of Eucalyptus largiflorens F.Muell. affected by short-term flooding. Aust J Ecol, 21(2): 173–183. |
| [] | Larcher W. 1997. Physiological plant ecology. Beijing: China Agriculture Press: 47–128. |
| [] | Mensforth L J, Thorburn P J, Tyerman S D, et al. 1994. Sources of water used by riparian Eucalyptus camaldulensis overlying highly saline groundwater. Oecologia, 100(1): 21–28. |
| [] | Mensforth L J, Walker G R. 1996. Root dynamics of Melaleuca halmaturorum in response to fluctuating saline groundwater, Plant and Soil, 184(1): 75-84. |
| [] | McCole A A, Stern L A. 2007. Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in Water. Journal of Hydrology, 342(2): 238–248. |
| [] | Peñuelas J, Filella I. 2003. Deuterium labelling of roots provides evidence of deep water access and hydraulic lift by Pinus nigra in a Mediterranean forest of NE Spain. Environmental and Experimental Botany, 49(2): 201–208. |
| [] | Querejeta J I, Estrada-Medina H, Allen M F, et al. 2006. Utilization of bedrock water by Brosimum alicastrum trees growing on shallow soil atop limestone in a dry tropical climate. Plant Soil, 287(2): 187–197. |
| [] | Querejeta J I, Estrada-Medina H, Allen M F, et al. 2007. Water source partitioning among trees growing on shallow karst soils in a seasonally dry tropical climate. Oecologia, 152(1): 26–36. DOI:10.1007/s00442-006-0629-3 |
| [] | Schwinning S, Ehleringer J R. 2001. Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. Journal of Ecology, 89(3): 464–480. DOI:10.1046/j.1365-2745.2001.00576.x |
| [] | Slavich P G, Smith K S, Tyerman S D, et al. 1999. Water use of grazed salt bush plantations with saline watertable. Agricultural Water Management, 39(1): 169–185. |
| [] | Smith S D, Wellington A B, Nachlinger J A, et al. 1991. Functional responses of riparian vegetation to streamflow diversions in the eastern Sierra Nevada. Ecol Appl, 1(1): 89–97. DOI:10.2307/1941850 |
| [] | Smith D M, Jarvis P G, Odongo J C W. 1997. Sources of water used by trees and millet in Sahelian windbreak systems. Journal of Hydrology, 198(1): 140–153. |
| [] | Sekiya N, Yano K. 2002. Water acquisition from rainfall and groundwater by legume crops developing deep rooting systems determined with stable hydrogen isotope compositions of xylem waters. Field Crops Research, 78(1): 133–139. |
| [] | Snyder K A, Williams D G. 2000. Watersources used by riparian trees varies among stream types on the San Pedro River, Arizona. Agricultural and Forest Meteorology, 105(1/3): 227–240. |
| [] | Socki R A, Karlsson H R, Gibson E K. 1992. Extraction technique for the determination of oxygen-18 in water using preevacuated glass vial. Analytical Chemistry, 64(6): 829–831. |
| [] | Schenk H J, Jackson R B. 2002. The global biogeography of roots. Ecol Monogr, 72(4): 311–328. |
| [] | Schenk H J, Jackson R B. 2005. Mapping the global distribution of deep roots in relation to climate and soil characteristics. Geoderma, 126(1): 129–140. |
| [] | Schwinning S. 2008. The water relations of two evergreen tree species in a karst savanna. Oecologia, 158(3): 373–383. DOI:10.1007/s00442-008-1147-2 |
| [] | Schwinning S, Starr B I, Ehleringer J R. 2005. Summer and winter drought in a cold desert ecosystem (Colorado Plateau) part Ⅰ: effects on soil water and plant water uptake. Journal of Arid Environments, 60(4): 547–566. DOI:10.1016/j.jaridenv.2004.07.003 |
| [] | Thorburn P J, Walker G R. 1993. The source of water transpired by Eucalyptus camaldulensis: soil, groundwater, or streams// Ehleringer J, Hall A, Farqubar G. Stable isotopes and plant carbon-water relations. San Diego:Academic Press, 511-527. |
| [] | Thorburn P J, Walker G R. 1994. Variations in stream water uptake by Eucalyptus camaldulensis with differing access to stream water. Oecologia, 100(2): 293–301. |
| [] | Thorburn P J, Ehleringer J R. 1995. Root water uptake of field-growing plants indicated by measurements of natural-abundance deuterium. Plant Soil, 177(2): 225–233. DOI:10.1007/BF00010129 |
| [] | Valentini R, Scarascia Mugnozza G E, Ehleringer J R. 1992. Hydrogen and carbon isotope ratios of selected species of a Mediterranean macchia ecosystem. Functional Ecology, 6(5): 627–631. |
| [] | Valentini R, Mugnozza G S, Angelis P D, et al. 1995. Coupling water sources and carbon metabolism of natural vegetation at integrated time and space scales, Agricultural and Forest Meteorology. 73(4/6): 297-306. |
| [] | Williams P W. 1985. Subcutaneous hydrology and the development of doline and cockpit karst. Z Geomorph N F, 29(4): 463–482. |
| [] | Williams P W. 1983. The role of the subcutaneous zone in karst hydrology. Journal of Hydrology, 61(1/3): 45–67. |
| [] | Walker C D, Richardon S B. 1991. The use of stable isotope of water in characterizing the source of water in vegetation. Chem Geol (Isotope Geosci Sect), 94(2): 145–158. DOI:10.1016/0168-9622(91)90007-J |
| [] | Williams D G, Ehleringer J R. 2000. Intra-and interspecific variation for summer precipitation use in pinyon-juniper woodlands. Ecological Monograph, 70(6): 517–537. |