 2012, Vol. 48
2012, Vol. 48文章信息
- 李兵兵, 秦琰, 刘亚茜, 刘相兵, 黄选瑞
- Li Bingbing, Qin Yan, Liu Yaxi, Liu Xiangbing, Huang Xuanrui
- 燕山山地油松人工林林隙大小对更新的影响
- Effects of Gap Size on Regeneration of Pinus tabulaeformis Plantation in the Yanshan Mountain
- 林业科学, 2012, 48(6): 147-151.
- Scientia Silvae Sinicae, 2012, 48(6): 147-151.
-
文章历史
- 收稿日期:2011-07-05
- 修回日期:2011-11-01
-
作者相关文章



2. 中国二十二冶集团第一建设有限公司 唐山 064000
2. The 1st Construction Corporation, Ltd of 22 MCC Group Tangshan 064000
林隙(gap)又称林窗,指森林群落中老龄树木死亡或偶然因素(如干旱、台风、火灾等)造成成熟阶段的优势种死亡,从而形成林冠空隙的现象(Watt,1947)。美国生态学家Runkle(1982)将林隙分为2种,一种是林冠空隙(canopy gap),指上方无树冠遮荫的部分,即太阳处于正中位置时无树冠遮荫的面积之和; 另一种是扩展林隙(expanded gap),指形成该林隙周围树木基部所包围的面积(Runkle,1981)。本研究所指的林隙为应用最为广泛的扩展林隙。近年来的森林动态学研究表明:林隙在森林结构和多样性维持中起着重要的作用,已成为当前森林生态学研究的最活跃领域之一(梁晓东等,2001a; 宋新章等,2006; 张春雨等,2006; 罗大庆等,2002)。林隙的形成,改善了林内光照条件,形成了不同于郁闭林分的异质性生境。林隙的大小能反映林隙环境与周围对照林分环境的差别程度,从而对林木种子萌发、苗木定居、幼树生长等自然更新过程产生重要影响(田悦颖,2007; 龙翠玲等,2007; 宋新章等,2007)。所以,林隙研究者常以林隙大小作为林隙干扰的重要特征之一,来研究树种的更新反应规律(Runkle,1981)。
我国在林隙方面的研究起步较晚,主要在海南岛热带雨林、温带阔叶红松林、亚热带常绿阔叶林、西南亚高山针叶林中进行了一些研究(宋新章等,2008a),研究对象多以天然林为主,而关于人工林人为干扰(如抚育间伐等措施)形成的林隙研究相对较少,间伐产生的林隙一般是小尺度和短暂的,但小尺度干扰对森林结构的发展,对林地状况如光、土壤含水量、凋落物分解等起到关键性作用(Myers et al., 2000),其重要性已成为国内外森林动态和天然更新研究中的共同论题(宋新章等,2008a; Zhu et al., 2003)。燕山山地分布着大面积的油松人工林,但由于各种因素的制约,多年来其林隙更新尤其是采伐林隙更新一直没有受到人们的注意。本文以此为研究对象,采用样线法进行野外调查,分析林隙大小对林冠下更新苗发生时间、更新密度、苗木质量以及草本多样性的影响,最终为人工油松单层同龄林诱导为复层异龄林提供依据。
1 研究区与林分概况 1.1 研究区概况研究区位于河北省木兰林管局境内的新丰林场。地处内蒙古高原和冀北山地的过渡带,地势西北高,东南低,海拔820~1 850 m,是阴山山脉、大兴安岭余脉向西南延伸和燕山余脉的结合部。属半干旱向半湿润过渡,寒温带向中温带过渡,大陆性季风型山地气候。无霜期67~125天,年平均气温-1.4~4 ℃,极端最低气温-42.9 ℃,极端最高气温38.9 ℃。年降水量380~560 mm,主要集中在7,8,9月。植被类型包括落叶针叶林、常绿针叶林、针阔叶混交林、阔叶林、灌丛或灌草丛、草原与草甸、沼泽及水生林分。乔木树种主要有油松(Pinus tabulaeformis)、华北落叶松(Larix principis-rupprechtii)、云杉(Picea asperata)、白桦(Betula platyphylla)、山杨(Populus davidiana)、蒙古栎(Quercus mongolica)、榆(Ulmus pumila)等; 灌木主要有榛子(Corylus heterophylla var. thunbergii)、沙棘(Hippophae rhamnoides)、胡枝子(Lespedeza bicolor)、土庄绣线菊(Spiraea pubescens)等; 草本植物主要有乌苏里苔草(Carex ussuriensis)、小花油点草(Tricyrtis macropoda)、歪头菜(Vicia unijuga)等。阴坡植被较好,盖度一般在30%~80%,阳坡次之,盖度一般在10%~20%。
1.2 林分概况研究对象为木兰林管局新丰林场南泉沟作业区210林班136小班的油松人工纯林,总面积6.68 hm2。造林时间为于1969年,采用2年生油松苗木,初植密度为4 995株·hm-2,期间经过6次人工抚育,分别于1984,1989,1999,2001,2004年和2006年进行。其中2006年结合大径材培育,进行了大强度人工抚育。目前林分平均密度为650株·hm-2,郁闭度为0.7,油松林冠下更新状况良好。
2 研究方法 2.1 调查方法1) 林隙特征调查本研究采用样线调查法,于2010年7—8月,在具有代表性的林分设置2条样线,样线长分别为150 m与180 m,对林隙进行调查。具体方法是,由坡下一随机点开始,沿着罗盘指的方向,遇到林隙时,实测起点到林隙距离,以所遇到的林隙中心为核心,记录样线中林隙的形状、长短轴以及边缘木的种类、胸径和高度。
2) 林隙内更新情况及多样性将所调查林隙按面积进行等级划分(以20 m2为区分段),选取各等级内具有代表性的林隙,调查其更新及多样性情况。
林隙更新调查在所调查的林隙中,统计各林隙内的更新苗总数; 利用手持罗盘及皮尺确定东西南北方向上的中心位置,在东西、南北方向上各设置一条宽为1 m的样带,2条样带的相交处设为中心样方记作C。E2,W2,S2,N2为东西南北边缘处的样方,E1,W1,S1,N1为C到4个方向边缘之间中心点上的样方,即每个林隙中9个1 m×1 m的小样方,样方内调查苗木的高及年龄。
草本生物多样性测定在林隙内十字交叉设置5个1 m×1 m的小样方,记录草本种名、株(丛)数、高度、盖度。
2.2 林隙面积的计算规则的近椭圆形的林隙面积,采用椭圆面积公式进行计算(Kneeshaw, 1998; 鲜骏仁等, 2004); 不规则且较大的林隙,采用Brokaw(1982)提出的等角多边形法计算,等角多边形法的原理是在林隙中心沿8或16个均分罗盘方向,将林隙近似看成等角8或16边形来估测林隙面积,采用形成木与边界木构建三角形,计算三角形面积,再求和来计算林隙面积(Brokaw, 1982)。
椭圆面积公式:A=πLW/4,
式中:A为椭圆形面积; L为椭圆长轴; W为椭圆短轴。
等角多边形法(海伦公式):A=[s(s-a)(s-b)(s-c)]/2,
式中:A为三角形面积; s为半周长; a, b, c为各边边长。
2.3 林隙更新密度差异性检验各林隙内更新苗木按高度区分为≤1.0 m的幼苗,1.0~2.0 m幼树,高度在2.0 m以上的小径木3个发育时期,计算各林隙内不同发育时期更新苗木的更新密度(更新密度=更新数量/林隙面积)。利用DPS 7.55软件进行最小显著差测试(least significant difference,LSD)。
2.4 物种多样性指数的计算物种多样性指数是群落的种类、个体总数以及各个种群个体的均匀程度的综合概念(李盾等,2004)。本研究利用ForStat软件对收集到的数据进行分析处理,计算Shannon-Wiener指数和Marglef丰富度指数。
Shannon-Wienner指数H的计算公式为:

|
Marglef丰富度指数R1计算公式:
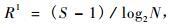
|
式中:S为物种数目; Pi为第i个物种在全部个体中的比例; N为群落中所有种的个体总数。
3 结果与分析 3.1 林隙大小特征林隙大小(gap size)是林隙的主要特征之一,通常用其面积来衡量,它是研究林隙环境和林隙更新最重要的一个指标(臧润国等,1998)。在所设置的样线中,共有45个林隙,其中面积最小的为20 m2,面积最大的为178 m2,把它们按面积大小进行等级划分(以20 m2为区分段),统计不同等级中林隙的数量。如图 1所示,林隙面积多在80 m2以下(数量比例为67.39%)。其中40~60 m2等级的所占数量比例和面积比例均为最大,分别为33.33%和22.22%。

|
图 1 林隙大小结构 Fig.1 Distribution of gaps sizes Ⅰ.20~40 m2; Ⅱ.40~60 m2; Ⅲ.60~80 m2; Ⅳ.80~100 m2; Ⅴ.100~120 m2; Ⅵ.120~140 m2; Ⅶ.140~160 m2; Ⅷ.160~180 m2 |
不同大小的林隙内,温度、水分、光照、土壤等生态因子及其组合会呈现不同特点,会对种子散播和苗木的生长产生不同效应,从而对林隙内的更新产生重要影响(梁晓东等,2001b; 臧润国等,1999; 臧润国,1998)。本研究以油松人工纯林为对象,更新的幼苗基本上都为油松苗,从苗木更新发生时间、苗木更新密度、苗木质量和草本多样性4个方面探讨了油松人工林林隙内的苗木天然更新规律。
1) 林隙大小与苗木更新发生时间的关系对不同大小林隙内更新苗的年龄进行调查分析发现,该地区林隙内的更新苗年龄主要为5~15年。以5年为一个等级(上线排外),将苗木分类。由表 1可知,更新苗年龄结构与林隙的面积有明显的相关关系。面积为20~100 m2的林隙,其苗木更新发生的时间较早,10~15年的更新苗数占所在林隙更新苗总数的80%以上; 面积为100~140 m2的次之,10~15年的更新苗数占所在林隙更新苗数的60%~68.02%;面积为140~160 m2的林隙,10~15年生更新苗数占所在林隙更新苗总数的46.08%;面积为160~180 m2的林隙最少,仅为26.67%。
|
|

2) 林隙大小对苗木更新密度的影响如表 2所示,处于不同发育时期的油松更新苗对林隙大小反应的趋势为:高度≤1.0 m的油松幼苗密度在林隙面积20~40 m2时最大,并且随着林隙面积的增大而减少,当林隙面积为160~180 m2时,更新密度最小,仅为475株·hm-2; 更新苗高度在1.0~2.0 m的幼树更新密度在面积为160~180 m2的林隙内最大,当面积为40~60 m2时,幼树的更新密度最小,为6 413株·hm-2; 高度≥2 m的小径木在林隙面积为20~40 m2时,更新密度最小,为2 761株·hm-2,在林隙面积为160~180 m2时,更新密度最大,为9 980株·hm-2。
|
|
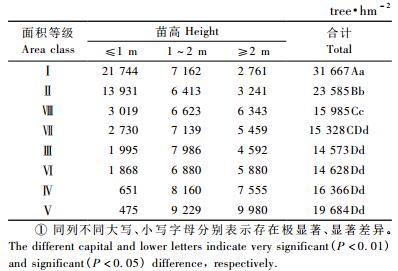
另外,由表 2可知,面积20~40 m2林隙不同发育时期油松更新苗(包括幼苗、幼树、小径木)密度的总和(31 667株·hm-2)极显著高于其他等级。这可能是由于小林隙单位面积下种量多的缘故,林隙越大,单位面积下种量越小,所以苗木密度小。随着林隙面积的增大,单位面积油松更新数量呈下降趋势。
3) 林隙大小对苗木质量的影响由表 1,2可知,高度≤1.0 m的油松幼苗更新密度在林隙面积20~40 m2时最大,而该等级林隙内年龄为10~15年的苗木占80.77%;高度≥2 m的油松更新苗密度在160~180 m2时最小,而该等级林隙中年龄为10~15年的苗木仅占26.67%。由此可以知道,小林隙更新苗时间早但高度低,而大林隙却恰恰相反。
4) 林隙大小对草本多样性的影响对林隙进行多样性调查,运用Shannon-Wienner指数反映各林隙内草本多样性,并利用Marglef公式计算丰富度指数,选择有代表性的林隙进行比较,分别建立林隙面积与Shannon-Wienner和Marglef丰富度指数的相关曲线。如图 2,3所示,草本植物的物种多样性、丰富度指数随林隙面积的增大,都呈现“单峰型”变化趋势,各最大值分别都出现在面积为60~80 m2的林隙。“单峰型”曲线呈左偏,说明中小林隙内的物种多样性要高于大林隙,反映出中小林隙较荫蔽的环境更适合燕山山地大多数草本种子的萌发、存活和生长。

|
图 2 林隙面积与Shannon-Wienner指数相关曲线 Fig.2 Relation curve between gap size and Shannon-Wienner index |

|
图 3 林隙面积与丰富度指数相关曲线 Fig.3 Relation curve between gap size and Marglef index |
林隙面积是衡量林隙干扰规模和强度的重要参数,也是决定林隙环境与非林隙环境差异的一个重要指标(龙翠玲,2008)。燕山山地油松人工林林隙面积范围为20~178 m2,以中小林隙为主,大林隙较少见,40~60 m2的林隙所占数量比例与面积比例均为最大。林隙以中小林隙为主,充分体现了该地区油松人工林隙多为人为干扰形成,但干扰强度较小的特点。由经营历可知,该地区曾经历过几次间伐,这是林隙形成的主要原因,但也不排除人为盗伐的因素。
不同大小的林隙对油松林隙更新的时间格局具有影响,较小的林隙苗木更新发生的时间较早,随着面积的增大,更新发生时间逐渐变晚,这是因为油松在幼苗阶段需要一定的荫蔽条件,面积较小的林隙不仅可以满足这一条件,而且为种子的发芽提供了适度的光照和温湿条件,同时可能还与种子雨有关,大林隙种子密度较小。
林隙大小对更新苗密度具有影响。龙翠玲(2008)对茂兰喀斯特森林林隙研究发现,单位面积树木幼苗和幼树个体数量(个体密度)和单位面积树木幼苗种数(种密度)随林隙面积增大而迅速减小。本研究也发现,面积为20~40 m2等级的林隙,其更新密度显著高于其他等级的林隙。林隙的存在,不但增加了光到达森林下层的持续时间,而且随着林隙面积的增加,林隙内的光照强度也明显增加,即光照强度是大林隙>小林隙>林下(彭建松等,2005)。油松属于阳性树种,因而在20~40 m2范围的林隙,表现出小径木最多的现象。相反,较大的林隙光照强烈,土壤干燥,灌草繁茂,不利于油松种子的萌发和出苗,即使林隙内有幼苗产生,也往往因灌木、杂草的竞争而不能正常生长,因此不利于幼苗幼树的生存,从而表现出,小林隙内幼苗、幼树更新密度高; 最大面积等级的林隙(160~180 m2)内幼苗、幼树更新密度高于小径木。
林隙大小对更新苗木质量影响显著。小林隙更新苗时间早但高度低,而大林隙却恰恰相反。小林隙一般主要由周围树木侧向生长来填充,而较大的林隙则主要靠更新树木向上生长来填充(宋新章等,2008b)。James等(2005)在热带雨林林隙中的更新试验表明:大林隙中幼苗的生长比小林隙明显快得多。在小林隙中,虽然苗木发生时间较早,但当苗木生长到一定时期,必然受到光照的限制,致使林隙内的更新苗年龄大苗高却很低; 而大林隙内的更新苗则主要是由于林隙的土壤干燥,种子不易萌发,幼苗也不易成活,但随着成活的苗木不断生长,林隙内的小气候也发生改变,已更新起的苗木为新生种子和幼苗提供了很好的庇荫作用,再加上光照较好,苗木普遍偏高,长势较好,形成了较为理想的异龄复层林分结构。
草本植物的物种多样性、丰富度指数随林隙面积的增大,都呈现“单峰型”变化趋势。“单峰型”曲线呈左偏,最大值都出现在面积为60~80 m2的林隙。说明中小林隙内的物种多样性要明显高于大林隙。这与林隙内生态因子(安树青等,1997),尤其是光因子的变化有关,太大或太小的林隙都不能拥有最大的物种多样性。当林隙面积小于80 m2时,随面积的增大曲线呈上升趋势,主要是因为林隙面积的增大为种子传播提供更大的空间,同时林隙内受到适当的光照,从而使种子得到使其发芽的有效积温。但随着林隙面积的增大,其环境条件的变化与中小林隙环境相比,差异较大,更接近于空旷地,光照强,土壤干燥不利于大多数种子的萌发和生存,因而其物种多样性也越低。
综上所述,林隙内的更新与林隙面积有着密切的关系。对于一些成熟林来说,可以人为制造林隙,为更新创造条件,林隙的面积开始应控制在20~40 m2较为合理,有助于林隙内更新苗的发生。当更新苗木长到一定时期(5年左右),应适当伐去上层木,扩大原有的林隙(80 m2左右为宜,此时的多样性及丰富度指数较高,群落结构较为稳定),为更新苗提供充足的光照,减少更新苗之间的相互竞争。苗木生长到10年左右,可采取适当的抚育措施(间苗等),所间下来的苗木可以作为大苗来销售。随后可继续扩大林隙,为保留下来的小径木提供广阔的生长空间,但最终保留的林隙面积应结合具体的经营目的来确定。这种人工促进天然更新的方法,不仅减少人力、物力,且能带来较大的经济效益,更能使林分形成较为稳定的异龄复层结构。
| [] | 安树青, 洪必恭, 李朝阳, 等. 1997. 紫金山次生林林窗植被和环境的研究. 应用生态学报, 8(3): 245–249. |
| [] | 李盾, 黄楠, 王强, 等. 2004. 天然次生林林木空间格局及更新格局. 东北林业大学学报, 32(5): 4–6. |
| [] | 梁晓东, 叶万辉. 2001a. 林窗研究进展(综述). 热带亚热带植物学报, 9(4): 355–364. |
| [] | 梁晓东, 叶万辉, 蚁伟民. 2001b. 林窗与生物多样性维持. 生态学杂志, 20(5): 64–68. |
| [] | 龙翠玲. 2008. 茂兰喀斯特森林林隙大小对树种更新的影响. 南京林业大学学报, 32(2): 35–39. |
| [] | 龙翠玲, 余世孝. 2007. 茂兰喀斯特森林林隙幼苗出现的时空格局. 云南植物研究, 29(5): 569–574. |
| [] | 罗大庆, 郭泉水, 薛会英, 等. 2002. 西藏色季拉山冷杉原始林林隙更新研究. 林业科学研究, 15(5): 564–569. |
| [] | 彭建松, 柴勇, 孟广涛. 2005. 云南金沙江流域云南松天然林林隙更新研究. 西北林学院学报, 20(2): 114–117. |
| [] | 宋新章, 李冬生, 肖文发, 等. 2007. 长白山区次生阔叶林采伐林隙更新研究. 林业科学研究, 20(3): 302–306. |
| [] | 宋新章, 肖文发. 2006. 林隙微生境及更新研究进展. 林业科学, 42(5): 114–119. |
| [] | 宋新章, 张智婷, 肖文发, 等. 2008a. 长白山杨桦次生林采伐林隙幼苗更新动态. 林业科学, 44(3): 13–20. |
| [] | 宋新章, 张智婷, 肖文发, 等. 2008b. 长白山次生杨桦林采伐林隙乔灌木幼苗更新比较研究. 林业科学研究, 21(3): 289–294. |
| [] | 田悦颖. 2007. 小兴安岭阔叶红松林林隙特征及其更新研究. 东北林业大学硕士学位论文. |
| [] | 鲜骏仁, 胡庭兴, 王开运, 等. 2004. 川西亚高山针叶林林窗特征的研究. 生态学杂志, 23(3): 6–10. |
| [] | 臧润国, 徐化成. 1998. 林隙(GAP)干扰研究进展. 林业科学, 34(1): 90–98. |
| [] | 臧润国, 余世孝, 刘静艳, 等. 1999. 海南霸王岭热带山地雨林林隙更新规律的研究. 生态学报, 19(2): 151–158. |
| [] | 臧润国. 1998. 长白山自然保护区阔叶红松林林隙更新的研究. 应用生态学报, 9(4): 349–353. |
| [] | 张春雨, 赵秀海, 郑景明. 2006. 长白山阔叶红松林林隙大小结构研究. 北京林业大学学报, 28(4): 34–38. |
| [] | Brokaw N V L. 1982. The definition of tree fall gap and its effect on measures of forest dynamics. Biotropica, 14(2): 158–160. DOI:10.2307/2387750 |
| [] | James G R, Matthew L, Mark S A, et al. 2005. Growth response by big-leaf mahogany (Swietenia macrophylla) advance seedling regeneration to overhead canopy release in southeast Para, Brazil. Forest Ecology and Management, 204(2/3): 399–412. |
| [] | Kneeshaw D D. 1998. Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology, 79(3): 783–794. DOI:10.1890/0012-9658(1998)079[0783:CGCATR]2.0.CO;2 |
| [] | Myers G P, Newton A C, Melgarejo O. 2000. The influence of canopy gapsize on natural regeneration of Brazil nut (Bertholletia excelsa) in Bolvia. For Ecol Man, 127(1/3): 119–128. |
| [] | Runkle J R. 1981. Gap regeneration in someold-growth forests of the eastern United States. Ecology, 62(4): 1041–1051. DOI:10.2307/1937003 |
| [] | Watt A S. 1947. Pattern and process in the plant community. Ecology, 35(1/2): 1–22. DOI:10.2307/2256497 |
| [] | Zhu J J, Lee F Q, Matsuzakit, et al. 2003. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. For Ecol Man, 182(1/3): 339–354. |