 2012, Vol. 48
2012, Vol. 48文章信息
- 张文辉, 杨文斌, 张洪, 陈美香, 宋启军
- Zhang Wenhui, Yang Wenbin, Zhang Hong, Chen Meixiang, Song Qijun
- 紫外光下毛竹颗粒的自由基与延迟发光
- Free Radical and Delayed Luminescence of Fibre of Moso Bamboo after UV-B Irradiation
- 林业科学, 2012, 48(6): 100-105.
- Scientia Silvae Sinicae, 2012, 48(6): 100-105.
-
文章历史
- 收稿日期:2011-05-03
- 修回日期:2011-08-23
-
作者相关文章


2. 福建农林大学材料工程学院 福州 350002;
3. 江南大学化学与材料工程学院 无锡 214122
2. College of Material Engineering, Fujian Agriculture and Forestry University Fuzhou 350002;
3. College of Chemistry and Material Engineering, Southern Yangtze University Wuxi 214122
毛竹(Phyllostachys edulis)既是一种生物材料,又是一种高分子聚合物,对其紫外光照射下的自由基和延迟发光的报道国内外尚属首次。自由基是由于分子在物理或化学因素的作用下,产生一个或一个以上的不配对电子,处于一种不稳定的高能级激发态。当这些电子从高能级激发态向基态跃迁时,能发出光子而发光。大多数自由基是通过各种物理与化学的方法把分子或基团中的共价键破坏,通过俘获或失去电子产生的(庞小峰,2007)。毛竹和木材一样,在紫外光的照射下将产生大量的自由基,同时,也产生了延迟发光(Keivanidis et al., 2010; Rothberg et al., 2008)现象。毛竹经机械刨切、粉碎后,断裂面的细胞壁被破坏,构成细胞壁的高聚物分子共价键断裂而产生机械自由基(Rex,1960; 许洪林等,1993)。毛竹经紫外光辐照后,高聚物表面裂解产生新的自由基。这些自由基有很高的表面活性,便于接枝反应的进行,能与胶合物质或涂料分子产生一系列化学反应形成新的连接,因此,增加毛竹表面自由基,可增加毛竹的表面自由能,有助于提高毛竹的胶合强度和涂饰效果(李坚,1994; 胡福增等,2001),对胶合板、刨花板、纤维板等的生产及毛竹表面的油漆、装饰均具有实用意义。本文将毛竹颗粒烘干到绝对含水率为零后,对绝干颗粒在紫外光照射下产生自由基和延迟发光进行数学拟合和物理意义的探讨,为最终将光信号转化成电信号、与计算机控制相结合、实现竹粉接枝共聚反应的自动控制探索了理论基础。
一个生物系统被光照射后的驰豫动力学过程中,生物光子辐射的相干性表现为辐射强度的非指数衰减,称为“延迟发光”(delayed luminescence)(顾樵,2007)。
1 材料与方法 1.1 设备德国Bruker公司产的型号为EMX-10/12的ESR电子自旋共振波谱仪; BPCL-1设备是由中国科学院生物物理研究所研制开发的,由微弱发光测量室、光电转换器、光子计数-脉冲放大电路、高稳定度高压电源、智能化数据测量与记录系统组成。样品室与光电倍增管相连, 信号被其接收后经光子放大器放大, 由数据采集系统收集后传送到计算机,样品室内有一个窗口和托架(其上放样品), 测量由样品室的快门控制。紫外灯是UV-B型灯管,功率20 W,波长302 nm,属于强效应波段,为生物有效辐射(郑有飞等,2009)。此外还有南京产DGH-9030型电热恒温干燥箱,上海第二天平仪器厂生产KS-016型分析天平。
1.2 材料采自福建省的3年生毛竹,取地上1~1.5 m的秆茎部分经过机械打磨成不同规格的深褐色竹粉,置于电热恒温干燥箱中60 ℃、8 h干燥至绝干后避光封存备用。
1.3 方法 1.3.1 毛竹自由基的测量在室温13 ℃下用分析天平称量样品80 mg放入样品池,在紫外灯下连续辐照60 min,每隔10 min利用ESR电子自旋共振波谱仪测一次样品自由基的波谱。毛竹自由基的测定用X波段100 kHz调制频率,用每克5×1014自由旋数的金刚石样品自由基强度换算毛竹试样每克自由旋数。由于本研究要求所得的自由基数据与延迟发光的数据做对比分析,因此,要求绝干毛竹颗粒的目数、辐照环境(温度、压力、紫外光的频率和强度)基本一致。毛竹受紫外辐照产生的自由基随时间增加而增长,因此,辐照不能断续进行,在紫外光辐照的同时进行自由基的电子自旋共振波谱(ESR)的测试。
1.3.2 毛竹延迟发光的测量在室温13 ℃下用分析天平称量样品0.592 g放入样品池,在紫外灯下辐照,每隔5 min利用BPCL设备测一次光强、动力学曲线和发射光谱。试验共使用10片窄带滤光片,10片滤波片的波长分别为400,425,440,460,490,535,555,575,620和640 nm,每换一片滤光片都进行同样的操作。测量光谱范围400~640 nm。测量参数为:工作电压1 000 V,测量时间60 s,间隔时间1 s,自动扣除本底。样品经紫外灯辐照后放入样品室开始记录。共测量3次,取平均值,误差在5%之内开始记录。
2 结果与讨论 2.1 毛竹自由基的波谱与紫外辐照时间的关系毛竹颗粒在紫外光连续辐照的同时,每隔10 min测1次ESR波谱,20目样品的波谱如图 1所示。

|
图 1 紫外光下毛竹的ESR波谱 Fig.1 ESR spectra of moso bamboo with UV-B irradiation a. 0 min; b. 30 min; c. 60 min. |
1) 辐照时间为0 min,即为未辐照的毛竹颗粒,在ESR波谱图上也看到自由基信号,这部分自由基称为机械自由基。
2) 毛竹经紫外光连续辐照后,其波谱与机械自由基波谱相比,线宽、线型未发生变化,峰高随辐照时间的增加而增加,光谱分裂因子g=2.003 3与机械自由基相同,在外磁场下,毛竹分子的自旋角动量起主导作用,轨道角动量不起作用; 紫外光辐照后的自由基强度比机械自由基明显增加。
2.2 毛竹自由基的强度与紫外辐照时间的关系不同规格毛竹颗粒自由基的绝对浓度随辐照能量的增加而快速增加,随时间延长而缓慢衰减,其关系如表 1所示。
|
|
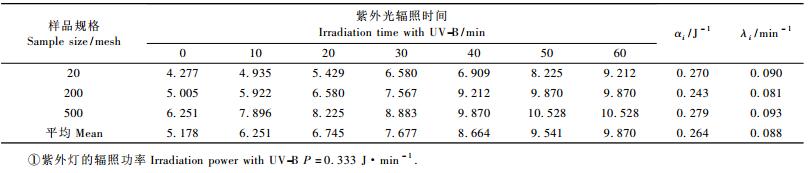
1) 毛竹机械自由基与比表面积有关,20目的最小,500目的最大。因为比表面积大的,单位质量的样品表面高聚物分子的共价健断裂数多,所以其机械自由基的数量就大。
2) 当毛竹颗粒所受紫外线辐照时间小于50 min时,自由基强度随辐照能量的增加而增加; 当辐照时间大于50 min时,自由基趋于饱和, 此时的自由基强度约为机械自由基的2倍。机械自由基强度大的500目毛竹颗粒, 受紫外光辐照相同时间和能量所产生的自由基强度也大。
3) 在紫外光辐照下,毛竹在产生新的自由基的同时,还有自由基的衰变。因此,毛竹中某种自由基随时间的变化关系可用下列微分方程表示:

|
(1) |
式中:P为紫外灯的功率; ηi为产生i种自由基的能量份额; εi为产生1个i种自由基所需能量; λa为i种自由基的衰变系数。
式(1)左边为任一时刻任意i种自由基变化率; 右边第1项为新自由基的产生率,第2项为该种自由基的衰变率,设衰变率与该种自由基数成正比。
把式(1)积分,并令t=0时,Ni=0,则:
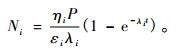
|
(2) |
由于紫外光辐照的时间在60 min内,毛竹样品吸收的能量可表示为:
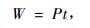
|
(3) |
代入式(2),得:
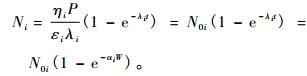
|
(4) |
式中:αi=λi/P,N0i=ηiP/εiλi=ηi/εiαi; Ni和N0i分别为i种自由基的瞬时值和最大值。
紫外光产生毛竹自由基的总和N为:
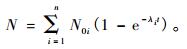
|
(5) |
从式(4),(5)可知,紫外光产生毛竹的某种自由基Ni按指数关系增长,但总自由基N一般不遵守这个规律。
把式(4)归一化,得:
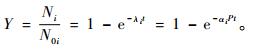
|
(6) |
当紫外光辐照的时间和总能量达到一定数量时,上式归一,即自由基趋于饱和。
4) 不同规格的毛竹颗粒紫外光辐照产生的自由基用式(6)进行回归,并作拟合曲线。20,200及500目的自由基浓度归一化后的理论和试验曲线如图 2所示。

|
图 2 毛竹紫外光产生自由基强度随辐照时间变化的理论与试验曲线 Fig.2 Theoretical and experimental curves of the variation of moso bamboo free radical after UV-B irradiation |
Y=1-e-αiPt所得的Y值是理论推导值,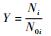
毛竹高聚物分子中存在大量的C—O和C—C键,这2种键的键能分别为3.7和3.6 eV。本研究采用302 nm处的紫外光,一个光子的能量是3.84 eV,它有足够的能量使高聚物分子键断裂而产生自由基。
由式(1)解得紫外光照产生的自由基在光照停止以后的衰变规律为:

|
(7) |
由于衰变系数λi很小,自由基的稳定性很好,自由基的稳定性源于毛竹碎裂成较小碎片或通过键断裂后进行重排的可能性很小,这与自由基的结构密切关联(Keivanidis et al., 2010)。毛竹聚合物在辐照分解过程中生成的自由基,由于位阻的困难,不能进行重结合反应,而是通过重排或歧化反应断链后被稳定下来。这就是毛竹受紫外光辐照后,产生比机械自由基大且稳定性好的自由基的机制。
2.4 毛竹延迟发光的光强与紫外辐照时间的关系 2.4.1 毛竹延迟发光的初始强度与紫外辐照时间的关系20目的毛竹颗粒在紫外灯下辐照,其延迟发光的初始光强每隔5 min紫外光照的测量值见表 2。样品从暗室中取出,未经紫外光辐照时没有延迟发光现象,初始光强为0;随着紫外光辐照时间的增加,延迟发光现象越来越显著,30 min时延迟发光的初始强度达到最强,为3 597,此时延迟发光持续60 s后光强降到940;紫外光辐照时间超过30 min后,样品的延迟发光现象变得更不明显,初始光强迅速减少,可能是紫外光辐照时间太长高分子聚合物样品出现老化所致(Kubera-Nowakowska et al., 2007)。毛竹样品的延迟发光在紫外光辐照30 min后出现饱和意味着生物光子辐照不是一个经典效应,而是一个典型的量子现象。
|
|
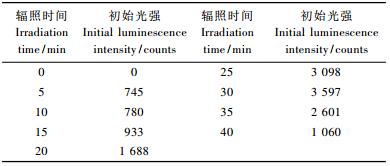
为了更好地研究毛竹颗粒延迟发光的衰减规律,将8条曲线分别叠加在2幅图上(图 3)。从图 3可以看出,30 min紫外光辐照时间的毛竹延迟发光强度最大,曲线下面的面积是总光强ΣI。生物系统的延迟发光一般遵从双曲线衰减,生物延迟发光的双曲衰减规律说明生物系统内各个激发态分子之间是相互偶联的,它们很可能是通过生物系统内相干的电磁场相互联系的。因此延迟发光可以作为一种新的研究方法用于研究竹材颗粒对外界环境的反应,并且延迟发光还可以反映生物内部的结构和相互作用的特性,从而使我们可以从系统角度理解和阐释生物生理状态的变化和机制。现代研究结果显示:延迟发光是非常敏感地依赖于许多环境因素和生理因素的(李光,2008)。

|
图 3 紫外光下毛竹颗粒的延迟发光曲线 Fig.3 Delayed luminescence curves of moso bamboo after UV-B irradiation |
为描述延迟发光所遵循的数学规律,从8条发光衰减曲线中选取4条(10,20,30和40 min)进行6种函数拟合研究,这6个函数分别是:
指数 I(t)=Ae-Bt; (1)
对数 I(t)=Alnt+B; (2)
幂指数 I(t)=At-B; (3)
多项式 I(t)=At2+Bt+C; (4)
双曲线1 I(t)=(A+Bt)-2; (5)
双曲线2 I(t)=(A+Bt)-1。 (6)
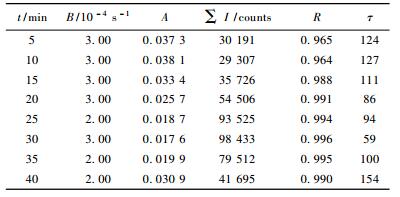
4条试验曲线经过多次拟合的最佳结果的相关系数如表 3所示。从表 3可以看出,指数、对数、幂指数、多项式与试验曲线的拟合结果差别很大,4个函数的相关系数R2相对双曲线都更小,说明毛竹的延迟发光也遵守双曲线衰减规律。式(5)和式(6)都是双曲线函数,相关系数R2都在0.96以上,2个公式在曲线变化规律上与试验数据吻合得较好。但总体而言,式(5)双曲线1的相关系数更好一些。
|
|

紫外光辐照刚结束,立即将毛竹样品放入暗室测量,得到的初始光强I0如表 2所示,它依赖于被测样品的性质,同时与辐照条件有关,延迟发光衰减的快慢与I0值的大小成正相关。式(5)I(t)=(A+Bt)-2反映延迟发光的各动力学参数的值,τ是相干时间(或衰减时间)(顾樵,2007),τ=A/B。当t→0时,I0=A-2,A=I0-1/2,紫外光辐照30 min时,I0最大,A最小,τ最小,延迟发光强度最大; 紫外光辐照5和10 min时,I0最小,A最大,τ也最大,延迟发光强度最小。显然A反映了延迟发光驰豫的初始速率,所以A是延迟发光初始光强的速率常数。
如果紫外光辐照样品结束后的足够长时间没有外界的物理、化学和生物的干扰,样品就不会发光了,当时t→∞,I(t)→0,说明数学公式与试验结果是吻合的,B是延迟衰减系数,与样品自身的性质有关。总光强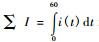
|
|
光谱是从一个介质中出来的光的强度与其波长的函数关系(Paras et al., 2006)。生物光子辐照存在于各种生物系统之中,光谱分布在可见光范围内是连续的。图 4是毛竹紫外光辐照30 min的光谱,其强度与波长的关系经过10片波片的测量显示:光谱分布在400~640 nm范围是连续的,620和640 nm的可见光出现峰值,分别占据延迟发光总强度的23.6%和59.5%。说明该波段内的光子的品质因子Q值小,相干性时间短,光子损耗大,延迟发光强,其他波段光子的品质因子相对较大,光子储存本领就大,因而延迟发光强度小。

|
图 4 紫外光辐照30 min毛竹的延迟发光强度随波长的变化 Fig.4 Variation of moso bamboo delayed luminescence intensity with change of wavelength |
紫外光下毛竹的自由基和延迟发光同时产生。自由基是一种高能激发态,极不稳定,衰减时以光子的形式辐射出去,构成延迟发光主要部分,毛竹延迟发光的能量一部分来自于自由基的衰减,另一部分来自于紫外光的光致发光,但这部分来源时间短,强度小。因此利用延迟发光的测量,就可以观察自由基的衰减情况,从而可间接测量毛竹的自由基浓度变化,为竹粉接枝共聚反应的自动控制提供理论依据。
3 结论1) 毛竹既是一种生物材料,又是一种高分子聚合物,绝干颗粒在紫外光照射下产生自由基的同时还产生延迟发光,自由基的光谱分裂因子g=2.003 3,自由基的强度随着辐照时间按Y=1-e-αiPt规律增加。
2) 毛竹自由基强度在紫外光辐照50 min时饱和,样品的延迟发光在紫外光辐照30 min出现饱和,紫外光辐照停止后,自由基强度按Ni=N0ie-λit的规律衰减,延迟发光按I(t)=(A+Bt)-2的规律衰减。自由基是一种高能激发态,它的稳定性是相对的,不稳定性是绝对的,延迟发光的能量一部分来自自由基的衰减,这部分是自由基发光,自由基和延迟发光有着内在的关系。
3) 延迟发光按双曲线规律衰减,公式中A是延迟发光的初始光强的速率常数,B是延迟衰减系数与样品自身性质有关,τ=A/B是相干时间,I是总光强。所有的延迟发光试验曲线和数学公式的相关系数R均大于0.96,故认为该公式在曲线变化规律上与试验吻合。
4) 光谱测量显示光谱分布在400~640 nm范围是连续的。620和640 nm的可见光出现峰值,说明该波段内的光子的品质因子Q值小,相干性时间短,光子损耗大,延迟发光强。
5) 研究自由基和延迟发光的产生和衰变规律,深入探讨拟合公式的物理意义,为最终将光信号转化成电信号、与计算机控制相结合、实现竹粉接枝共聚反应的自动控制探索了理论基础。
| [] | 顾樵. 2007. 生物光子学. 北京, 科学出版社. |
| [] | 胡福增, 郑安呐, 张群安. 2001. 聚合物及其复合材料的表界面. 北京, 中国轻工业出版社. |
| [] | 李光. 2008. 生物超微落发光的非线形分析. 河北大学学报:自然科学版, 28(2): 142–145. |
| [] | 李坚. 1994. 木材科学. 哈尔滨, 东北林业大学出版社. |
| [] | 庞小峰. 2007. 生物物理学. 成都, 电子科技大学出版社. |
| [] | 许洪林, 张文辉. 1993. 木材表面机械自由基的研究. 南京林业大学学报, 17(1): 69. |
| [] | 郑有飞, 吴荣军. 2009. 紫外辐射变化及其作物响应. 北京, 气象出版社. |
| [] | Keivanidis P E, Kamm V, Dyer-Smith C, et al. 2010. Delayed luminescence spectroscopy of organic photovoltaic binary blend films: probing the emissive non-geminate charge recombination. Advanced Materials, 22(45): 5183–5187. DOI:10.1002/adma.201002389 |
| [] | Kubera-Nowakowska L, Lichszteld K, Kruk I. Delayed luminescence of solid polymers. Rev Adv Mater Sci, 14: 90–96. |
| [] | Paras N. 2006. 生物光子学导论. 何赛灵, 译. 杭州: 浙江大学出版社, 85. |
| [] | Rex R W. 1960. Electron paramoneuc resonance studies of stable free radicals in lignins and humic acide. Nature, 188(4757): 1185–1186. DOI:10.1038/1881185a0 |
| [] | Rothberg L J, Chen S H, Cuppoletti C, et al. 2008. Studies of delayed luminescence in conjugated polymers, 235th ACS National Meeting. New Orleans, LA, United States: April 6–10, 1008. |