 2012, Vol. 48
2012, Vol. 48文章信息
- 张雁, 郭同斌, 诸葛强
- Zhang Yan, Guo Tongbin, Zhuge Qiang
- 转Bt基因南林895杨对杨扇舟蛾体内酶的影响
- Effects of Bt Nanlin 895 Poplar on the Metabolic Enzymes of Scarce Chocolate Tip, Clostera anachoreta
- 林业科学, 2012, 48(6): 95-99.
- Scientia Silvae Sinicae, 2012, 48(6): 95-99.
-
文章历史
- 收稿日期:2012-03-15
- 修回日期:2012-04-08
-
作者相关文章



2. 徐州市森林病虫防治检疫站 徐州 221004
2. Forest Pest Management and Quarantine Station of Xuzhou Xuzhou 22100
害虫摄入杀虫剂以后,体内的酶尤其是与解毒有关的酶活性会发生相应变化,以缓解毒害。昆虫解毒酶是一类异质酶系,能够代谢大量的内源或外源有毒物质从而维持正常的生理代谢,解毒酶可通过改变自身特性增强解毒能力,其体内相关代谢酶的变异是导致害虫产生抗药性的重要机制之一(Hilder et al., 1987)。这些解毒酶一般包括酯酶(esterase)、羧酸酯酶(carboxylesterase,CarE)、多功能氧化酶(mix-function oxidase,MFO)和谷胱甘肽S-转移酶(glutathione S-transfer,GST)等(翟启慧等,1995),而乙酰胆碱酯酶(acetylcholinesterase,AChE)是神经突触部位清除乙酰胆碱、维护神经正常传导的重要酶类,并且许多杀虫剂都以其作为靶标来实现杀虫效果,因而它与昆虫的解毒功能有关(李飞等,2003)。孙芸芹等(1987)发现棉蚜(Aphis gossypii)的解毒代谢抗性与酯酶及羧酸酯酶活力密切相关。朱鹏飞等(1992)研究发现杀虫剂溴氢菊酯有效抑制了敏感品系的马尾松毛虫幼虫(Dendrolimus punctata punctata)的酯酶活性,而抗性品系幼虫的酯酶活性变化幅度则较小。陈松等(2002)研究认为棉铃虫(Helicoverpa armigera)体内酯酶活性的升高与抗药性有关。除了上述这些酶类之外,在逆境条件下昆虫体内的保护酶即超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(catalase,CAT)和过氧化物酶(peroxidase,POD)产生协调作用而处于动态平衡状态,以防止逆境产生的自由基的伤害(Wang et al., 2001),所以这些保护酶的活性也与昆虫耐药性、抗逆性有关。郭同斌等(2006)研究发现杨小舟蛾取食转Bt单价及转Bt+CpTI双价基因杨树叶片后,其体内的CAT和POD活性受到抑制,而SOD活性却有所上升。
南林895杨(Populus deltoides×P. euramericana‘Nanlin 895’)是南京林业大学从美洲黑杨与欧美杨杂交后代群体中选育出的优良无性系,它适应范围广、稳定性强、生长快、材质好,可作纤维板和纸浆用材以及农田林网和四旁植树等绿化树种,目前已在黄淮、江淮及长江中下游等地区广泛种植。诸葛强等(2006)利用农杆菌介导法转化Bt基因获得转Bt基因南林895杨,试验表明对靶标害虫有明显的抑制作用。已有研究表明:转Bt基因杨树对美国白蛾(Hyphantria cunea)、杨小舟蛾(Micromelalopha sieversi)的中肠酯酶和乙酰胆酯酶存在一定的抑制作用(郭同斌等,2007),但目前尚缺乏对主要害虫杨扇舟蛾的解毒酶及保护酶体系的系统研究。笔者于2010年7—9月间以转Bt基因南林895杨当年生扦插苗为材料,系统测定杨扇蛾幼虫取食转Bt基因杨树叶片后体内解毒酶和保护酶的变化,研究了转Bt基因杨树对害虫生理生化的影响,为防治害虫的应用及抗性治理提供参考依据。
1 材料与方法 1.1 植物材料及虫源转Bt基因南林895杨树株系B21及未转化的对照(CK)南林895杨,均来自南京林业大学林木遗传和基因工程实验室。2009年4月底从林间采集杨扇舟蛾,经室内用对照杨树叶饲养幼虫至3龄时作为试验虫源。取个体大小一致、刚蜕皮的3龄幼虫,饥饿2 h后用株系B21植株的顶部第3,4片展开叶饲喂至4,12,24,36,48 h后解剖截取中肠。对照(CK)杨作同样处理。
1.2 酶源制备在每个时间点处理计10头幼虫,5次重复共50头作为提取中肠的虫源。将10头幼虫置于预冷的蜡盘中截取中肠,-20 ℃冰箱中冰冻贮存。测前取出稍融,加入410 mL的0.02 mol·L-1磷酸盐缓冲液匀浆,离心取上清液作为酶源。
1.3 乙酰胆碱酯酶的测定参考Gorun(1978)改进的Elman的方法。取0.1 mol·L-1、pH 7.4的磷酸缓冲液2.4 mL,酶源0.2 mL;取Asch-DTNB(1:2)混合液0.1 mL水浴中振荡,恒温15 min,然后加1×10-3 mol·L-1毒扁豆碱0.5 mL;取0.1 mol·L-1磷酸缓冲液2.6 mL,取Asch-DTNB(1:2)混合液0.1 mL振荡水浴中25 ℃,恒温15 min;然后加1×10-3 mol·L-1毒扁豆碱0.5 mL作对照,在7200型分光光度计412 nm处测定。据酶源蛋白质含量计算乙酰胆碱酯酶活力。
1.4 谷胱甘肽S-转移酶的测定取酶液0.2 mL,66 mmol·L-1pH 7.4的磷酸缓冲液2.4 mL;50 mmol·L-1GSH 0.3 mL;以0.03 mol·L-1CDNB 0.1 mL作对照;取66 mmol·L-1 pH 7.0的磷酸缓冲液2.6 mL;50 mmol·L-1GSH 0.3 mL;以0.03 mol·L-1CDNB 0.1 mL作对照。将各试剂立即混匀,于25 ℃、340 nm下测定5 min内OD变化值。据酶源蛋白含量,计算谷胱甘肽S-转移酶比活力(nmol·mg-1 min-1)。每处理重复3次。
1.5 酯酶和羧酸酯酶的活力测定酯酶测定参照李飞等(2003)的方法。取150 μL固牢兰RR盐和α-醋酸萘酯混合液,50 μL 0.02 mol·L-1、pH 7.0的磷酸盐缓冲液,并加入25 μL酶液,置于酶标仪中检测,25 ℃,450 nm。每处理重复3次。多功能氧化酶参照丁双阳等(2001)的方法。
1.6 超氧化物歧化酶的测定按改进的Beauchamp(1971)和Fridovich(1977)方法测定。反应孔中含50 mmol·L-1磷酸盐缓冲液(pH 7.0),13 mmol·L-1蛋氨酸,75 μmol·L-1氮蓝四唑,0.1 mmol·L-1乙二胺四乙酸及10 μL酶液、4 μmol·L-1核黄素。置于4 000 lx日光下进行光化学反应,25 ℃,15 min后黑暗终止反应,立即在550 nm处比色。一个酶活单位相当于引起300 μL反应液达到50%抑制所需酶量。
1.7 过氧化氢酶活性测定按改进的程鲁京等(1994)的方法。酶促反应后剩余的H2O2与钼酸铵形成稳定的黄色复合物,其颜色深浅与酶活性成反比。在酶标板的空白孔B1中加入100 μL基液(65 μmol·mL-1 H2O2),100 μL钼酸铵,20 μL酶液。空白孔B2中加入100 μL基液,100 μL钼酸铵,20 μL pH 7.4磷酸盐缓冲液。测定孔加入20 μL酶液,100 μL基液,37 ℃保温1 min后,立即加入100 μL钼酸铵摇匀,于405 nm处测光密度值。每分钟分解1 μmol过氧化氢为一个酶活力单位。
1.8 过氧化物酶活性测定按改进的Simon等(1974)的方法。反应孔中含100 mmol·L-1磷酸盐缓冲液(pH 6.0),30 mmol·L-1愈伤木酚,26 mmol·L-1 H2O2。饲喂转Bt基因及对照杨树叶片时间处理为4,12,24 h的中肠酶源加样量为20 μL,36,48 h为5 μL,作用5 min后用于470 nm处测光密度值。酶活单位以每毫克蛋白每分钟在470 nm处的OD值表示。
1.9 酶源蛋白质含量测定采用考马斯亮蓝G2250染色法测定各处理的酶源蛋白质含量。
2 结果与分析 2.1 转Bt基因南林895杨对酯酶和羧酸酯酶的影响在杨扇舟蛾取食转Bt基因杨树叶片4~48 h内,酯酶活性即开始持续下降(表 1)。与至4 h时相比,至24 h时酯酶活性下降了32%,至36 h时下降了57%,至48 h时下降了62%。从36 h后酯酶活性与对照(CK)相比,差异性显著(P < 0.05),至48 h时达到抑制高峰,与对照(CK)相比,差异性极显著(P < 0.05)(表 1)。
|
|
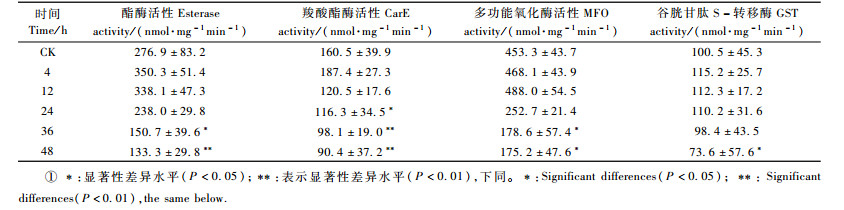
羧酸酯酶活性在害虫取食前期的4 h时,与对照CK的平均水平相比有所上升,可能跟害虫体内自我保护的应激反应有关,但从12 h后其酶活性受到抑制并持续下降,与CK相比,24 h后即出现显著性差异,至36 h时差异极显著(P < 0.01)。说明转Bt基因杨树对酯酶和羧酸酯酶有明显抑制效果,随着饲喂时间的增加,抑制效果增大。
2.2 对多功能氧化酶、谷胱甘肽S-转移酶和乙酰胆碱酶的影响多功能氧化酶在害虫取食叶片的4~12 h内活力高于CK的平均值并且呈现递增趋势,但从24 h起,其活性受到抑制,此后呈下降趋势,并与对照相比有显著性差异。谷胱甘肽S-转移酶在前36 h内波动很小,但在取食48 h后与4 h时相比下降了36%并且与CK相比差异性显著(P < 0.05)(表 1)。乙酰胆碱酶的活力在4~12 h内呈上升态,12 h后呈缓慢下降趋势,但与对照相比无显著性差异,说明Bt毒蛋白对其虽有影响但并不剧烈,这可能与Bt毒蛋白不是神经性毒素有关。
2.3 对害虫体内相关保护酶的影响超氧化物歧化酶活性在害虫体内前12 h内高于照CK,虽略有升高但变化不大,然而在害虫取食转Bt基因叶片的12~48 h内受到了Bt毒蛋白的抑制,其活性持续下降。取食转Bt基因叶片至36 h时的酶活性与4 h时相比下降了近一半,与对照CK相比,36 h后出现显著性差异(P < 0.05)(表 2)。而与之相反,过氧化物酶活性水平却在4~48 h内呈持续上升趋势,害虫取食转Bt基因叶片24 h后与对照相比,酶活性出现显著性差异(P < 0.05),48 h后差异极显著(P < 0.01)(表 2)。过氧化氢酶活性在4~12 h时段呈下降态,24~48 h时段内呈上升态,至48 h时与对照呈显著差异性(P < 0.05)。
|
|

酯酶和羧酸酯酶是昆虫的重要解毒酶系,有不少科学工作者先后探讨了其对外源有毒物质的应答规律。丁双阳等(2001)研究发现转基因杨树对美国白蛾幼虫中肠的酯酶和羧酸酯酶有明显抑制作用,且其活性随着害虫取食时间的增加而迅速下降;与之不同,张少燕等(2004)用含不同浓度Bt毒蛋白的人工饲料饲喂棉铃虫,发现随着Bt毒蛋白浓度的提高,幼虫体内的羧酸酯酶活力呈上升趋势;郭同斌等(2007)研究发现杨小舟蛾取食转基因杨树叶片后,体内酯酶和羧酸酯酶在初期皆先呈上升趋势然后呈下降趋势。张炬红等(2011)研究发现转双抗虫基因741杨显著抑制了羧酸酯酶活性。本研究表明靶标害虫杨扇舟蛾体内的中肠酯酶和羧酸酯酶与上述研究不尽相同:中肠酯酶从一开始即受到转基因杨Bt毒蛋白持续抑制,这方面与丁双阳等(2001)的研究一致,而羧酸酯酶的活性却从进食转基因叶片初期有所上升以适应外界毒素的影响,但随后即受到持续抑制,这方面与郭同斌等(2007)的研究结果类似。总的来说Bt毒蛋白对这2种酶都有较明显的抑制作用,通过抑制害虫中肠酯酶和羧酸酯酶的活力而干扰其正常生理代谢,从而导致靶标害虫发育缓慢或死亡。虽然多功能氧化酶和谷胱甘肽S-转移酶在代谢外源底物中起到重要作用,但本研究发现转Bt基因杨对它们的活性虽有抑制但效果不突出,到害虫进食36 h及48 h后才有明显下降迹象,其原因还有待进一步研究。乙酰胆碱酯酶是神经突触部位清除乙酰胆碱、维护神经正常传导的重要酶类,它的活性关系到昆虫神经生理活性的正常与否。本研究发现转Bt基因杨树对此酶的抑制作用并不显著,饲喂害虫至48 h时才有活性下降表现,这可能与Bt毒蛋白不属于直接的神经性毒素有关。
超氧化歧化酶能清除害虫体内O2·生成H2O2,H2O2又能与O2·形成毒性更强的HO·,此时过氧化氢酶和过氧化物酶负责分解H2O2防止产生更多的自由基伤害虫体。Govenor等(1997)研究舞毒蛾(Lymantria dispar)幼虫取食含有外源毒素物质的槐树(Sophora japonica)叶片后,中肠超氧化歧化酶活性升高,过氧化氢酶活性上升;蒋继宏等(2005)用苦豆碱处理杨小舟蛾幼虫后,其体内过氧化氢酶活性和过氧化物酶活性下降,超氧化歧化酶在前期上升;郭同斌等(2006)发现杨小舟蛾在取食转Bt和CpTI双抗虫基因的杨树叶片后,超氧化物歧化酶活性上升,其他2种保护酶下降。本研究发现:超氧化物歧化酶仅在初期略有上升,但不具备显著差异性(P < 0.05),饲喂害虫24h以后其活性呈下降趋势,而其他2种酶在后期呈明显的上升状态。这种情况的产生可能由于Bt毒蛋白扰乱了虫体保护酶系统,使其在后期超氧化物歧化酶活性受阻,降低了清理O2·能力,从而生成了更多的H2O2导致虫体产生适应性反应,使得另外2种保护酶的活性上升以阻止产生更多的自由基,当然也可能与杨扇舟蛾幼虫对Bt毒白特异性应答有关。
| [] | 陈松, 杨亦桦, 吴益东. 2002. 不同种群棉铃虫三龄幼虫酯酶活性频率分布与抗药性的关系. 昆虫学报, 45(6): 733–738. |
| [] | 程鲁京, 孟泽. 1994. 钼酸铵显色法测定血清过氧化氢酶. 临床检验杂志, 12(1): 6–8. |
| [] | 丁双阳, 李怀业, 李学锋, 等. 2001. 转Bt基因杨树对美国白蛾幼虫中肠解毒酶及乙酰胆碱酯酶的影响. 东北林业大学学报, 29(3): 28–30. |
| [] | 郭同斌, 嵇保中, 蒋继宏, 等. 2006. 转基因杨树对杨小舟蛾体内三种保护酶活力的影响. 昆虫学报, 49(3): 381–386. DOI:10.7679/j.issn.2095-1353.2006.094 |
| [] | 郭同斌, 嵇保中, 诸葛强, 等. 2007. 转基因杨树对杨小舟蛾幼虫解毒酶活性的影响. 林业科学, 43(5): 59–63. |
| [] | 蒋继宏, 吴薇, 曹小迎, 等. 2005. 苦豆碱对杨小舟蛾体内保护酶系统活力的影响. 南京林业大学学报, 29(5): 91–93. |
| [] | 李飞, 韩召军, 唐波. 2003. 抗性品系棉蚜乙酰胆碱酯酶和羧酸酯酶的变异. 昆虫学报, 46(5): 578–583. |
| [] | 孙芸芹, 冯国蕾, 袁家畦, 等. 1987. 棉蚜对有机磷杀虫剂抗性的生化机理. 昆虫学报, 30(1): 13–20. |
| [] | 翟启慧. 1995. 昆虫分子生物学的一些进展:杀虫剂抗性的分子基础. 昆虫学报, 38(4): 493–501. |
| [] | 张炬红, 高宝嘉, 刘军侠. 2011. 转双抗虫基因741杨对靶标害虫主要解毒酶和乙酰胆碱酯酶的影响. 东北农业大学学报, 42(1): 66–70. |
| [] | 张少燕, 李典谟, 谢宝瑜. 2004. Bt毒蛋白对棉铃虫的生长发育和相关酶活性的影响. 昆虫知识, 41(6): 536–540. DOI:10.7679/j.issn.2095-1353.2004.136 |
| [] | 朱鹏飞, 王荫长, 尤子平. 1992. 溴氢菊酯对马尾松毛虫幼虫血淋巴和中肠酯酶的影响. 林业科学研究, 5(2): 188–192. |
| [] | 诸葛强, 房丹, 李秀芬, 等. 2006. 美洲黑杨杂种优良无性系转抗虫基因(Bt和CpTI)的研究. 分子植物育种, 4(6): 819–824. |
| [] | Beauchamp C D, Fridovich I. 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analytical Biochemistry, 44(1): 276–287. DOI:10.1016/0003-2697(71)90370-8 |
| [] | Fridovich I. 1977. Oxygen is toxic!. Bioscience, 27(7): 462. DOI:10.2307/1297527 |
| [] | Gorun V, Proonov L. 1978. Modified Ellman procedure for assay of cholinesterase in crude enzymatic preparation. Analytical Biochemistry, 186: 324–326. |
| [] | Govenor H L, Schultz J C, Appel H M. 1997. Impact of dietary allelochemicals on Gypsy moth (Lymantria dispar) caterpillars: Importance of midgut alkalinity. Journal of Insect Physiology, 43(12): 1169–1175. DOI:10.1016/S0022-1910(97)00049-8 |
| [] | Hilder V A, Gatehouse A M R, Sheerman S E, et al. 1987. A novel mechanism of insect resistance engineered into to tobacco. Nature, 330: 160–163. DOI:10.1038/330160a0 |
| [] | Simon L M, Fatrai Z, Jonas D E, et al. 1974. Study of peroxide metabolism enzymes during the development of Phaseolus vulgris. Biochemistry Physiology, 166(5/6): 388–392. |
| [] | Wang Y, Oberley L W, Murhammer D W. 2001. Evidence of oxidative stress following the viral infection of two lepidopteran insect cell lines. Free Radical Biology and Medicine, 31(11): 1448–1445. DOI:10.1016/S0891-5849(01)00728-6 |