 2012, Vol. 48
2012, Vol. 48文章信息
- 李洁, 姚延梼, 周春娥, 梁志英, 苗青
- Li Jie, Yao Yantao, Zhou Chun'e, Liang Zhiying, Miao Qing
- 分区交替水分胁迫对草乌光合特性的影响
- Effect of Alternate Partial Root Zone Drying on Photosynthesis Characteristics of Aconitum kusnezoffii
- 林业科学, 2012, 48(6): 72-79.
- Scientia Silvae Sinicae, 2012, 48(6): 72-79.
-
文章历史
- 收稿日期:2011-12-26
- 修回日期:2012-04-01
-
作者相关文章



2. 河南师范大学生命科学学院 新乡 453007;
3. 晋中学院生命科学院 晋中 030600
2. College of Life Sciences, Henan Normal University Xinxiang 453007;
3. College of Life Sciences, Jinzhong College Jinzhong 030600
草乌(Aconitum kusnezoffii),为毛茛科(Ranunculaceae)乌头属多年生植物,可作为林下资源用于退耕还林(草)工程,花呈蓝紫色,是宝贵的蓝色花卉基因资源和良好的观花植物,其块根可入药,也是重要的濒危药用植物。近些年来的掠夺式挖取使其野生资源濒临枯竭,人工栽培前途广阔,但目前我国干旱缺水趋势日趋严重,给草乌的正常生长和推广应用带来了严峻考验。有关乌头属植物的研究大都集中在资源调查(周先建等,2011)、药用价值(刘文龙等,2011)和栽培技术(魏书琴,2011)、组织培养(张继等,2010)等方面,关于其抗旱性研究很少,对其在干旱条件下的综合开发有一定的盲目性。光合作用是植物基本的生命活动,也是对逆境影响最为敏感的生理过程之一,逆境胁迫对植物光合特性的影响程度决定着植物的抗性水平和产量形成,可用作判断植物生长和抗逆性强弱的指标(谭永芹等,2010)。
根系分区交替水分胁迫方法源于控制性交替灌水技术,该技术使根系某个区域保持干燥,而让一部分区域灌水湿润,交替使不同区域的根系经受一定程度的水分胁迫锻炼,刺激根系吸收补偿功能,以利于部分根系处于水分胁迫时产生的根源信号脱落酸(ABA)传输至地上部叶片,调节气孔开度,达到以不牺牲作物光合产物积累而大量减少其蒸腾耗水,从而产生节水效应(康绍忠等,2001)。目前该技术已成为植物生理科学和灌溉科学研究的热点(Tahi et al., 2007;Topcu et al., 2006;Davies et al., 2002)。Tiago等(2003)曾证明分根区交替灌溉可使葡萄(Vitis vinifera)水分利用率提高80%,而产量基本相当;程福厚等(2007)证明分区灌溉可以使梨树(Pyrs spp.)总水分利用率较传统灌溉提高22.9%,果实产量和品质无明显差别;Davies等(2002)对柑桔(Citrus reticulata)、橄榄(Canarium album)、柚子(Citrus maxima)、西红柿(Lycopersicum esculentum)、草莓(Fragaria daltoniana)、苹果(Malus pumila)等植物的研究表明:交替灌溉可以大量节水,产量保持同等水平或略有下降,但可促进糖分向果实转移,提高口味和品质(Davies et al., 2000)。但该技术在一些密植类作物中很难达到,普适性还需进一步提高,而稀植类作物中应用该技术时也需进一步简化操作流程,提高简易性(柴强等,2010)。
本文以人工栽培的草乌为研究对象,在田间应用控制性交替灌水技术原理设计水分胁迫,研究根系分区交替水分胁迫(alternate partial root zone drying,APRD)、固定单侧水分胁迫(fixed partial root zone drying,FPRD)与对照两侧同时供水(both partial root zone irrigation,BPRI)对草乌光合特性的影响,旨在研究抗旱机制的同时为草乌在干旱地区的综合开发利用提供参考,并验证根系分区交替供水的节水效应,为其进一步在田间推广应用提供可操作的方法和依据。
1 试验地概况试验在山西农业大学苗圃进行,112°53′E,37°30′ N,海拔870 m,年降水量540 mm,无霜期为140~180天。试验地土壤质地为壤土,pH 7.85,有机质含量9.22 g ·kg-1,土壤氮、磷、钾含量分别为1.1 g ·kg-1,11.6 mg ·kg-1,115.7 mg ·kg-1。
2 材料与方法 2.1 试验材料所用材料为2年生草乌实生苗,草乌种子采自于山西省关帝山,2009年4月中旬播种于苗床,幼苗出土后生长缓慢,秋季枯萎,于结冻前在苗床上铺一层腐殖土越冬。2010年4月按株距30 cm、行距60 cm垄植移栽,待6月份长出4~5片叶片后开始水分胁迫处理。
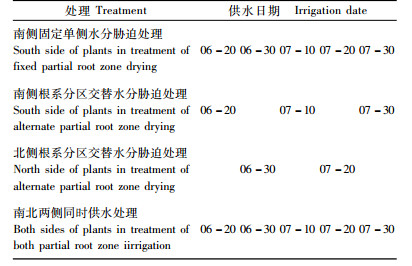
2.2 试验方法APRD为分区交替水分胁迫,6月20日先给植株一侧(南侧)供水,10天后给另一侧供水,此时先供水侧不供水,继续胁迫,20天后再次给先供水侧供水,后供水侧继续胁迫至30天时再次供水,如此循环进行“供水—胁迫—再供水”过程(即每10天分侧供水1次,每侧供水周期间隔为20天,期间随着土壤水分自然消耗逐渐产生水分胁迫),供水量参照先期相关研究(李洁等,2011),每次单侧供水30 mm;FPRD为固定单侧水分胁迫,即每10天给植株固定的一侧(南侧)供水1次,每次供水30 mm;BPRI为对照,每10天给植株两侧同时供水1次,每次每侧供水30 mm。试验从第1次给APRD处理植株南侧供水开始共经历3个周期,具体供水安排见表 1。
|
|
为减少土壤水分侧向渗漏,在距植株10 cm处开梯形沟(沟为东西方向),沟底宽10 cm,沟深30 cm,用软管进行灌水,控制灌水量。每个处理30株,设置3个重复,完全随机区组排列,设置保护行和遮雨棚,遮雨棚只在降雨时使用。试验期间除灌水外,各处理的其他田间管理均保持一致。
2.3 指标测定 2.3.1 土壤含水量测定在2次供水间隔的中间及下次供水的前1天和供水当天,下午19:00左右,在每个处理的每个重复中分散选取5株,在植株的南北两侧分别用土钻按每20 cm为一层分层进行取样,取至根系80 cm处,装入铝盒中,带回实验室称鲜质量,在105 ℃烘至恒质量,计算土壤质量含水量,即水分质量占干土质量的百分比。
2.3.2 光合特性测定光合日变化测定在每个周期末晴天(具体测定日期为7月8日,7月29日,8月17日,其中第3次测定日期因故调整),在每个处理的每个重复内,选择5株长势较一致的植株,选其中上部生长健壮的成熟叶片标记3片,用便携式光合作用测定系统(Li-6400,Li-COR)进行田间连体测定,叶片夹好后将叶室垂直对着太阳光,测定时间为7:00—18:00,每1 h测定1次,包括光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、水分利用效率(WUE),WUE用单叶瞬时净光合速率和蒸腾速率之比表示(宋锋惠等,2010):WUE=Pn/Tr。每个处理对应取15次重复所测平均值,绘制各光合指标的日变化曲线。
选择光照强度、温度及湿度相对稳定的3个晴天,9:00—11:30,测定不同处理下的光合-光响应过程,红蓝光源设置的光合有效辐射为1 400,1 200,1 000,900,800,600,400,300,250,200,150,100,50,0 μmol·m-2s-1,每梯度持续3 min,其余方法与日变化测定相同,设置叶温为28 ℃,CO2浓度约360 μmol·mol-1,为开放气路,相对湿度为40%~60%。
2.3.3 光合-光响应曲线拟合采用分段函数方法(段爱国等,2010)将弱光阶段与强光阶段分别拟合得到光合-光响应曲线,分段函数表达式如下:
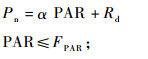
|
(1) |

|
(2) |
式中,FPAR为弱光与强光分界点处的光强(本研究中FPAR为200 μmol·m-2s-1),参数α为直线段的斜率,即植物表观光合量子效率(AQY),Rd为暗呼吸速率。由(1)式可知光补偿点LCP(Pn为0时的PAR),由-Rd /α计算可得。光饱和点(LSP)和最大净光合速率(Pmax)根据实测数据点的走势估计得出。
2.4 统计分析采用Microsoft Excel 2003和SPSS17.0统计分析软件进行数据处理,用Duncan法进行处理间0.05水平的差异显著性检验。
3 结果与分析 3.1 不同水分处理对土壤含水量的影响图 1为各处理植株南北两侧80 cm深度内的土壤水分动态变化。FPRD北侧为未灌水侧,其土壤含水量较低,基本呈持续缓慢下降趋势,其南侧土壤含水量再次供水后迅速恢复,且随着每轮周期的进行逐渐下降;APRD北侧第1次所示结果为该侧供水前的土壤含水量7.56%,此时南侧土壤含水量为15.12%,每对一侧进行供水后,另一侧土壤含水量稍有波动;对照BPRI植株南北两侧的土壤含水量变化趋势十分接近。

|
图 1 不同水分胁迫下植株南北两侧土壤含水量变化 Fig.1 The changes of soil water content of plant two sides under different water stress FPRD,APRD,BPRI分别代表固定单侧水分胁迫处理、根系分区交替水分胁迫处理、两侧同时供水处理FPRD,APRD and BPRI represented the treatment of fixed partial root zone drying, alternate partial root zone drying and both partial root zone irrigation respectively.下同。The same below. |
图 2为草乌在3种水分条件下的Pn-PAR响应曲线,由图 2A可知:在光强小于200 μmol·m-2s-1的弱光区,草乌叶片的Pn随光强增强以不同斜率直线式的升高。草乌在3种水分条件下的光响应过程基本相似,在800 μmol·m-2s-1光强以内,Pn表现为以递减的增速随光强上升,之后FPRD、对照BPRI在光强为1 000 μmol·m-2s-1左右时趋于饱和,APRD处理在光强为1 250 μmol·m-2s-1时趋于饱和。由于从不饱和到饱和是1个渐进过程,没有明显的转折点,因此饱和光强是1个范围,而不是1个点(许大全,2002),此处饱和光强取光强范围的上限。测定时的光合有效辐射日变化如图 3所示。

|
图 2 不同水分处理下草乌的Pn-PAR响应曲线 Fig.2 The response curve of Pn to PAR of Aconitum kusnezoffii under different water treatments |

|
图 3 光合有效辐射日变化 Fig.3 Diurnal changes of PAR |
将3种水分条件下Pn和PAR进行二元回归可得模拟方程(2)和弱光区的直线段方程(1)(表 2),各方程的决定系数均大于0.95,说明拟合曲线合理,较好地反映了所设条件下叶片对光的响应过程。通过对表 2和图 2进行分析计算,得到草乌在3种水分条件下的光合-光响应特征参数(表 3)。
|
|
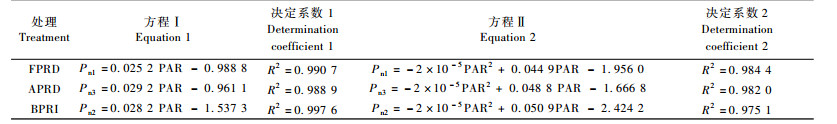
|
|
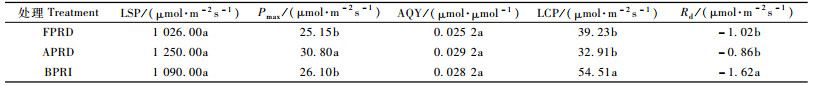
AQY反映植物对光能的利用效率和光合物质生产效率(段爱国等,2010)。从图 2和表 3可知:3种水分条件下AQY表现为APRD>对照BPRI>FPRD,但各处理间差异不显著。在交替水分胁迫下草乌的光能利用率有所提高,但仍低于植物最大量子效率的理论值0.08~0.125 μmol·μmol-1(Long et al.,1994)。
Rd反映植物在没有光照条件下的呼吸速率,与叶片的生理活性有关(伍维模等,2007)。草乌的Rd表现为APRD处理稍低于FPRD处理,二者均显著低于对照BPRI,说明在此条件下草乌叶片呼吸减弱,在一定程度上减少光合产物的消耗,以抵御对干物质积累的影响。
3.2.2 对光饱和点(LSP)和光补偿点(LCP)的影响LSP和LCP分别代表植物利用强光与弱光的能力,3种水分条件下草乌利用强光的能力没有显著差异;APRD处理下LCP最小,对弱光的利用能力有所增强,其次为FPRD处理,二者均显著低于对照BPRI。APRD处理使草乌LCP降低的同时,其LSP有所提高,此协同变化表明根系分区交替水分胁迫在一定程度上增大了草乌对光环境的适应范围。由表 3可知:草乌的最大光强为1 250 μmol·m-2s-1,依有关分类方法属阳生植物(孟繁静,2000),但在强光条件下会发生光抑制,在推广栽培时应考虑此因素,林药结合模式较为合理。
3.2.3 对最大净光合速率(Pmax)的影响在一定生境下,Pmax反映了植物叶片的最大光合能力(Iryna et al., 2004)。本试验中,Pmax表现为APRD>对照BPRI>FPRD,APRD处理下潜在光合能力显著高于FPRD和对照BPRI。对照BPRI耗水量大,水分胁迫较少或程度较轻没有形成相应的逆境生理调节机制,在遇到干旱时其最大光合能力较易受抑制;FPRD处理虽然供水也减少50%,但由于未供水侧根系长期处于水分亏缺状态,使其光合能力下降。由试验结果可知:土壤水分状况和供水方式对草乌光合能力有显著影响,APRD处理具有明显优势。
3.3 不同水分处理对草乌光合日变化的影响 3.3.1 对叶片Pn日变化的影响对照BPRI Pn日变化在不同测定日均呈双峰曲线,峰值出现在10:00, 15:00,随着“供水-胁迫”的循环,日均值减小(图 4)。第1轮中,FPRD, APRD处理下Pn呈单峰曲线,峰值分别出现在10:00,11:00,日均值表现为:对照BPRI>FPRD>APRD。第2轮各处理Pn变化较大,FPRD处理下Pn呈非典型单峰曲线;APRD处理下Pn呈双峰曲线,上午峰值出现在9:00,比对照提前,下午峰值出现在15:00,日均值表现为:对照BPRI>APRD>FPRD。第3轮FPRD处理下Pn呈单峰曲线,峰值出现在10:00;APRD处理下Pn呈双峰曲线,上午峰值出现最晚(11:00),此后Pn开始大于对照BPRI,下午15:00达第2次高峰,其日均值(10.58 μmol·m-2s-1)比对照BPRI, FPRD处理分别提高4.16%, 37.43%,交替水分胁迫下草乌通过自身生理生化调节其Pn逐渐高于对照,这有利于干物质积累。

|
图 4 不同水分处理下草乌叶片Pn日变化 Fig.4 Diurnal changes of photosynthetic rate(Pn)of leaves of Aconitum kusnezoffii plants under different water condition A,B,C分别代表 3个周期中的测定日,7月8日、7月29日、8月17日A,B and C represented the test date in the first,second and third cycles respectively,July 8th,July 29th,August 17th.下同。The same below. |
Gs降低或气孔关闭是植物抵御干旱胁迫的第一道防线(Sitza et al., 2004)。3种水分条件下Gs日变化除第3轮FPRD处理下午峰值不明显外,基本均表现为双峰曲线,上午峰值出现在9:0,下午峰值出现在15:0或16:0(图 5)。对照BPRI的Gs高于APRD,FPRD处理。第1,3轮时,APRD处理Gs日均值最低,第2轮时FPRD处理Gs日均值最低,三者差异较大。由图 4可知:对照BPRI Gs最高,而其Pn在后期却低于APRD处理,这是因为影响Gs的主要环境因子除了土壤水分亏缺还包括空气湿度,而影响光合作用的因素除了气孔因素外,也包括非气孔因素,所以二者虽然关系密切,但不一定同步变化或互为直接因果。

|
图 5 不同水分处理下草乌叶片Gs日变化 Fig.5 Diurnal changes of stomatal conductance(Gs)of leaves of A.kusnezoffii plants under different water condition |
3种水分条件下,草乌叶片Tr日变化基本呈单峰曲线,随着循环的进行,日均值均逐渐降低(图 6)。对照BPRI Tr一直高于APRD, FPRD处理,第1, 2轮峰值出现在10:00,第3轮峰较平缓;前2轮中,APRD处理与FPRD处理差异较小;第3轮中,FPRD处理下Tr降幅较大,随着水分亏缺的加重,峰值减小,峰出现时间前移。就3次测定的总均值比较,APRD处理Tr稍高于FPRD处理,比对照BPRI下降30%,而其Pn却高于对照BPRI,反映出交替水分胁迫对草乌气孔行为的优化调节。

|
图 6 不同水分处理下草乌叶片Tr日变化 Fig.6 Diurnal changes of transpiration rate(Tr)of leaves of Aconitum kusnezoffii plants under different water condition |
WUE是评价水分亏缺条件下植物生长适宜程度的一个综合生理生态指标(王颖等,2006)。3种水分条件下WUE从清晨开始逐渐增大,高峰在8:00—11:00出现,之后随着Pn的降低和Tr的升高逐渐下降,傍晚时有1次升高(图 7)。第1轮时,对照BPRI的WUE日均值最高,APRD与FPRD处理相近;第2轮时,对照BPRI与FPRD处理WUE下降,APRD处理WUE上升,日均值表现为APRD>对照BPRI>FPRD;第3轮时,FPRD与APRD处理WUE上升幅度较大,表现为APRD>FPRD>对照BPRI。随着土壤水分的不断减少,各处理WUE逐渐升高,从总均值来看,APRD处理下WUE分别比对照BPRI和FPRD处理提高20.39%和10.53%,而其供水量只有对照BPRI的1/2,这对草乌在水资源紧缺的干旱地区的开发利用有重要意义。

|
图 7 不同水分处理下草乌叶片WUE日变化 Fig.7 Diurnal changes of water use efficiency(WUE)of leaves of A.kusnezoffii plants under different water condition |
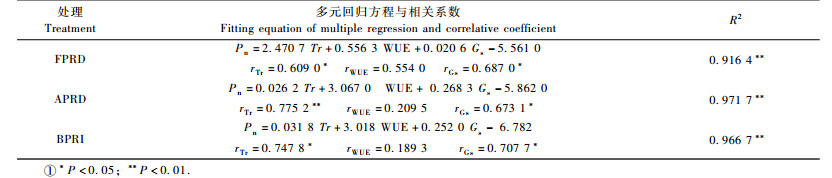
从表 4可知:3种水分条件下Pn与Tr, WUE, Gs的回归模型R2均大于0.90(F < 0.01),该模型可以合理反映其回归关系。APRD处理下Tr的回归系数最小(0.026 2),在Tr下降相同量的情况下,其Pn下降幅度最小,其次是对照BPRI,FPRD降幅最大,这也是经历一定阶段循环后APRD处理在Tr降低的条件下Pn却较高的原因之一;3种水分条件下Gs与Pn的回归表现正好与Tr相反,Pn还受非气孔因素的影响。从相关系数来看,在FPRD,APRD,对照BPRI条件下,与Pn相关性最大的因素分别是Gs(0.687 0), Tr(0.775 2), Tr(0.747 8),均达到显著或极显著正相关。WUE在FPRD处理下与Pn的相关性远大于APRD处理与对照BPRI,说明随着土壤水分亏缺的加重,WUE与光合作用之间相关性增大,但均未达显著水平。
|
|
在干旱和半干旱地区,水分不足是限制植物光合作用的一个因素,但植物进行光合作用时作为原料消耗的水只是从土壤中吸收的水的很小一部分,其余绝大部分都是通过蒸腾作用散失掉的,因此,水分亏缺对Pn的影响并不是由于原料水供应不足,而是由于水分亏缺引起的气孔或非气孔的限制(许大全,2002)。气孔是CO2和水汽交换的通道,气孔行为同时控制着叶片的光合与蒸腾,蒸腾作用对气孔有较强的依赖性,而光合作用除受气孔因素影响外,还受非气孔因素的影响(Giacomo et al., 2005)。因此适当降低Gs可以大大降低蒸腾耗水,而对光合作用却可以影响很小或没有影响(康绍忠等,2001)。本试验中,经过一定阶段的“供水-胁迫”循环,APRD处理在供水量减少一半的条件下,Tr比对照降低30%,而Pn高于对照4.16%;FPRD处理虽然供水量也减少一半,Tr低于对照和APRD处理,但其Pn大大降低,结合3种水分条件下Gs表现为对照BPRI>FPRD>APRD可知,APRD处理可优化调节气孔行为,在非充分灌溉条件下可实现水分的优化分配。此外,在APRD处理下Pn得以提高也与草乌反复经历“供水-胁迫”所产生的旱后补偿生长效应有关,这与安玉艳等(2010)对杠柳(Periploca sepium)的研究结果相似。而草乌作为以块根入药的植物,其地上部叶片的光合作用将会影响到地下部分块根的大小和内含物的积累。
4.2 根系分区交替水分胁迫对草乌光合-光响应的影响水分胁迫可直接引发光合机构的异常,同时也影响与暗反应有关的酶活性及光合电子传递,而Pmax决定于Rubisco活性和电子传递速率(Walting et al., 2000)。本研究中,草乌的光合-光响应曲线拟合结果表明:在试验条件下草乌的光合机构可正常运转。经过一定阶段的根系分区交替水分胁迫,草乌的Pmax显著提高,AQY有所增大,光适应性增强,Rd显著降低,与有关研究结论一致(Chaves et al., 2004;Flexas et al., 2004;Ennahli et al., 2005)。但这一结果是由于PSⅡ电子传递速率和叶肉细胞Rubisco活性变化所致,还是由于其中某一因素或其他因素影响所致,有待进一步研究。
本试验中,草乌的AQY介于0.025~0.03 μmol·μmol-1之间,低于在适宜生长条件下测定的一般植物的AQY范围0.03~0.05 μmol·μmol-1(李合生,2002),这可能与本试验所设置的水分胁迫条件有关,同时不同的光合光响应模拟模型所产生的结果也有一定差异;也有一些植物在适宜生长条件下,其AQY低于0.03 μmol·μmol-1(张昆等,2009)。经过水分胁迫后,草乌Rd有不同程度的降低,这一结果与韩刚等(2010)对柠条(Caragana korshinskii)的相关研究结论一致,但与柯世省等(2007)对夏腊梅(Sinocalycanthus chinensis)的相关研究结论相反;根系分区交替水分胁迫可降低草乌的LCP,与左应梅等(2011)对木薯(Manihot esculenta)的相关结论一致,但与韩刚等(2010)及柯世省等(2007)的相关结论有一定差异。这可能与植物自身对水分胁迫的敏感程度和抗性不同有关,同时也与各研究者所设置的试验条件和胁迫标准的不同有关。
该试验设计水分胁迫时所采用的根系分区供水方法主要为草乌在干旱地区的田间栽培提供依据和方法,与以往室内营养液控制模式或盆栽模式下的分根灌水技术相比,精确度和控制性不如后者,但更接近于实际生产。对于在田间如何采取更合理的根系分区供水方法,有研究者进行了探索。常见方法为隔行灌溉;杜太生等(2006)曾对葡萄进行隔沟交替灌溉,每次灌水后有一定侧向渗漏,但2~3天内即趋于稳定;也有研究者采用双管交替滴灌的方式(周青云等,2011),精确度较好,但投资成本增加,且在密植类作物上难以应用。经本试验证明,该方法可以产生节水的效果,且操作流程简化,普适性较高。干湿交替不仅节约灌溉水,还能平衡营养生长与生殖生长的矛盾,促进光合产物向经济产量转化(William et al., 2000)。例如,干湿交替可以使葡萄和葡萄酒品质(杜太生等,2006)以及其他一些作物品质大大改善(杨建昌等,2005),关于根系分区交替水分胁迫对草乌地下部块根产量和品质产生的影响有待进一步研究。
| [] | 安玉艳, 郝文芳, 龚春梅, 等. 2010. 干旱-复水处理对杠柳幼苗光合作用及活性氧代谢的影响. 应用生态学报, 21(12): 3047–3055. |
| [] | 柴强. 2010. 分根交替灌溉技术的研究进展与展望. 中国农业科技导报, 12(1): 46–51. |
| [] | 程福厚, 赵志军, 张纪英, 等. 2007. 分区交替灌溉对梨生长结果及水分利用效率的影响. 干旱地区农业研究, 25(4): 130–133. |
| [] | 杜太生, 康绍忠, 张霁, 等. 2006. 不同沟灌模式对沙漠绿洲区葡萄生长和水分利用的效应. 应用生态学报, 17(5): 805–810. |
| [] | 段爱国, 张建国, 何彩云, 等. 2010. 干热河谷主要植被恢复树种干季光合光响应生理参数. 林业科学, 46(3): 68–73. DOI:10.11707/j.1001-7488.20100311 |
| [] | 韩刚, 赵忠. 2010. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 30(15): 4019–4026. |
| [] | 康绍忠, 潘英华, 石培泽, 等. 2001. 控制性作物根系分区交替灌溉的理论与试验. 水利学报(11): 80–86. DOI:10.3321/j.issn:0559-9350.2001.11.014 |
| [] | 柯世省, 金泽新. 2007. 干旱胁迫和复水对夏蜡梅幼苗光合生理特性的影响. 植物营养与肥料学报, 13(6): 1166–1172. DOI:10.11674/zwyf.2007.0629 |
| [] | 李合生. 2002. 现代植物生理学. 北京, 高等教育出版社: 125. |
| [] | 李洁, 周春娥, 梁志英, 等. 2011. 干旱胁迫对乌头部分生理指标的影响. 水土保持科学, 9(3): 93–100. |
| [] | 刘文龙, 刘志强, 宋凤瑞, 等. 2011. 乌头类双酯型生物碱组分转化为单酯水解型及脂型生物碱组分的研究. 高等学校化学学报, 32(3): 717–720. |
| [] | 孟繁静. 2000. 植物生理学. 武汉, 华中科技大学出版社: 206-213. |
| [] | 宋锋惠, 罗青红, 史彦江, 等. 2010. 平欧杂交榛4个品种光合特性的比较研究. 新疆农业科学, 47(5): 876–881. DOI:10.6048/j.issn.1001-4330.2010.05.006 |
| [] | 王颖, 魏国印, 张志强, 等. 2006. 7种园林树种光合参数及水分利用效率的研究. 河北农业大学学报, 29(6): 44–48. |
| [] | 谭永芹, 柏新富, 朱建军, 等. 2010. 渗盐分与水分胁迫对三角叶滨藜和玉米光合作用的影响. 生态学杂志, 29(5): 881–886. |
| [] | 魏书琴. 2011. 黄花乌头根腐病菌的生物学特性研究. 北方园艺(1): 169–171. |
| [] | 伍维模, 李志军, 罗青红, 等. 2007. 土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响. 林业科学, 43(5): 30–35. |
| [] | 许大全. 2002. 光合作用效率. 上海, 上海科学技术出版社: 39. |
| [] | 杨建昌, 袁莉民, 唐成, 等. 2005. 结实期干湿交替灌溉对稻米品质及籽粒中一些酶活性的影响. 作物学报, 31(8): 1052–1057. |
| [] | 张继, 田玉汝, 杨宁, 等. 2010. 野生高乌头组织培养及快速繁殖. 生物学通报, 45(1): 40–41. |
| [] | 张昆, 万勇善, 刘风珍, 等. 2009. 花生幼苗光合特性对弱光的响应. 应用生态学报, 20(12): 2989–2995. |
| [] | 周青云, 王仰仁, 孙书洪. 2011. 根系分区交替滴灌条件下葡萄根系分布特征及生长动态. 农业机械学报, 42(9): 59–63. |
| [] | 周先建, 夏燕莉, 张美, 等. 2011. 四川省野生乌头资源调查研究. 资源开发与市场, 27(5): 448–449. |
| [] | 左应梅, 杨重法, 唐建, 等. 2011. '华南8号'木薯光合特性对土壤水分响应的研究. 中国农学通报, 27(3): 107–111. |
| [] | Chaves M M, Oliveira M M. 2004. Mechanisms underlying plant resilience to water deficits:prospects for water-saving agriculture. Journal of Experimental Botany, 55(407): 2365–2384. DOI:10.1093/jxb/erh269 |
| [] | Davies W J, Bacon M A, Thompson D S, et al. 2000. Regulation of leaf and fruit growth in plants growing in drying soil: exploitation of the plants' chemical signalling system and hydraulic architecture to increase the efficiency of water use in agriculture. Experimental Botany, 51(350): 1617–1626. DOI:10.1093/jexbot/51.350.1617 |
| [] | Ennahli S, Earl H J. 2005. Physiological limitations to photosynthetic carbon assimilation in cotton under water stress. Crop Science, 45(6): 2374–2382. DOI:10.2135/cropsci2005.0147 |
| [] | Flexas J, Bota J, Loreto F, et al. 2004. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biology, 6(3): 269–279. DOI:10.1055/s-2004-820867 |
| [] | Iryna I T, Michael M B. 2004. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remobilization of apple leaves. New Phytologist, 164(3): 441–450. DOI:10.1111/j.1469-8137.2004.01197.x |
| [] | Long S P, Humphries S, Falkowski P G. 1994. Photo inhibition of photosynthesis in nature. Annu Rev Plant Physiol Mol Biol, 45(1): 633–662. DOI:10.1146/annurev.pp.45.060194.003221 |
| [] | Sitza A G, Killingbeck K T, Whitford W G. 2004. Plant water relations and photosynthesis during and after drought in a Chihuahuan desert arroyo. Journal of Arid Environments, 59(1): 27–39. DOI:10.1016/j.jaridenv.2004.01.011 |
| [] | Tiago P S, Carlos M L, Lucilia M R, et al. 2003. Partial root zone drying effects on growth and fruit quality of field-grown grapevines(Vitis vinifera). Func Plant Biol, 30(6): 663–671. DOI:10.1071/FP02180 |
| [] | Tahi H, Wahbi S, Warkrim R, et al. 2007. Water relations, photosynthesis, growth and water use efficiency in tomato plants subjected to partial rootzone drying and regulated deficit irrigation. Plant Biosyst, 141(2): 265–274. DOI:10.1080/11263500701401927 |
| [] | Topcu S, Kirda C, Dasgan Y. 2006. Yield response an N-fertilizer recovery of tomato grown under deficit irrigation. European Journal of Agronomy, 26(1): 64–70. |
| [] | Walting J R, Press M C, Quick W P. 2000. Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum. Plant Physiology, 123(3): 1143–1152. DOI:10.1104/pp.123.3.1143 |