 2012, Vol. 48
2012, Vol. 48文章信息
- 尹吴, 李丽莎, 王立科, 王明庥, 诸葛强
- Yin Wu, Li Lisha, Wang Like, Wang Mingxiu, Zhuge Qiang
- 转玉米PEPC基因杨树的光合生理特性分析
- Analysis of Photosynthetic Characteristics of Transgenic Poplars with Maize PEPC Gene
- 林业科学, 2012, 48(6): 63-71.
- Scientia Silvae Sinicae, 2012, 48(6): 63-71.
-
文章历史
- 收稿日期:2011-07-27
- 修回日期:2012-04-17
-
作者相关文章



根据光合作用途径的不同,可将植物分为C3,C4和CAM植物。与C3植物相比, C4植物由于具有CO2浓缩机制,能在高光强、高温及低CO2浓度、干旱等条件下有较高的光合效率及营养和水分利用效率(Moore,1982;Ku et al., 1996)。随着分子生物学的兴起和植物转基因技术的快速发展,许多与C4光合作用途径相关的关键酶如磷酸烯醇式丙酮酸羧化酶(PEPC)、NADP-苹果酸酶(NADP-ME)和丙酮酸磷酸双激酶(PPDK)等一系列基因已从玉米(Zea mays)、高粱(Sorghum vulgare)和苋菜(Acalypha)等C4植物中克隆获得,且已有许多关于C4植物光合相关基因导入C3植物进行高光效育种的报道(焦德茂等,2001;李霞等,2001;Edgerton,2009;Bar-Even et al., 2010;Berg et al., 2010)。
杨树(Populus)(C3植物)分布广、种类多,是重要的速生用材树种和绿化造林树种之一。近年来,我国杨树人工林面积已达700万hm2,位居世界第一,对我国乃至世界的环境保护和气候变化都起到了重要作用。本文首次在实验室利用转基因技术获得的一批转玉米PEPC基因杨树的基础上,以未转基因南林895杨(Populus deltoides×P. euramericana ‘Nanlin895’)为对照,转PEPC基因杨树的光合生理特性分析,为杨树高光效分子育种提供试验依据。
1 材料与方法 1.1 试验材料以南林895杨为材料, 经农杆菌介导法,将玉米PEPC基因(PEPC基因表达载体由香港中文大学生物学教授、中国工程院院士Samuel Sai-ming SUN教授惠赠)导入南林895杨中,共获得50个株系,通过PCR检测分析及生长初步筛选,从15个检测为阳性的株系中选择10个转基因株系与未转基因的南林895杨同时盆栽于温室,每个株系4~5盆,每盆栽1苗,常规管理,经常喷水以保持环境湿度。
1.2 光合作用相关酶的测定光合作用相关酶活测定于7,8月进行,根据光合作用相关酶的氧化还原反应,在340 nm处测定光吸收值的变化计算酶活性。酶活性测定的前提是反应时间和酶浓度呈线性关系。用紫外可见光分光光度计(UV-4802型)分析测定光合作用相关的酶如PEPC,PPDK,NADP-ME,NADP-MDH和RuBPc的活性(Kung et al., 1980;Chen et al., 1981;Gonzalez et al., 1984;Li et al., 1987;Wei et al., 1994;Ku et al., 1999;Olcer et al., 2001;Sage,2002;Portis et al., 2007)。
1.3 光合速率相关参数的测定光合速率相关参数的测定于8月进行,采用GFS-3000便携式气体交换-荧光测定仪(GFS-3000)测定转PEPC基因杨树与对照植株的光合速率及相关参数。
1.3.1 光合速率(Pn)的日变化在晴朗无云天气,7:00—19:00每隔1 h测定光合速率日变化,测定时采用树冠中部外围着生于枝条顶端的3~5对叶片,每株共测定3片,每次测定记录5次。
1.3.2 蒸腾速率(Tr)与胞间CO2浓度(Ci)的日变化测定过程基本与光合速率(Pn)的日变化一致。
1.3.3 光能利用效率(LUE)与水分利用效率(WUE)的日变化通常用净光合速率(Pn)与蒸腾速率(Tr)的比值来表示瞬间水分利用效率(郑淑霞等,2006);用净光合速率(Pn)与光合有效辐射(PAR)的比值来表示光能利用效率(马成仓等,2004)。测定过程与光合速率(Pn)日变化的测定基本一致。
1.3.4 光合作用对光的响应测定时间为上午10:00—11:30。将光合有效辐射在0~1 500 μmol·m-2s-1范围内,设定若干梯度测定光合速率。测定时通过系统控制叶室温度为30 ℃,叶室CO2浓度为350 μmol·mol-1 ,相对湿度60%。光饱和点、光补偿点、表观量子效率、暗呼吸速率以及光饱和时的光合速率等参数:利用光合作用对光响应的相关数据,通过回归分析获得。
1.3.5 光合作用对CO2的响应通过光合测定系统调节CO2浓度,光合速率在0~1 000 μmol·mol-1范围内设定若干梯度测定,测定时叶室温度为30 ℃,光合有效辐射为1 200 μmol·m-2s-1,相对湿度60%。CO2饱和点、CO2补偿点、羧化效率、光呼吸速率、CO2饱和时的光合速率等参数,利用光合作用对CO2响应的有关数据,通过回归分析获得。
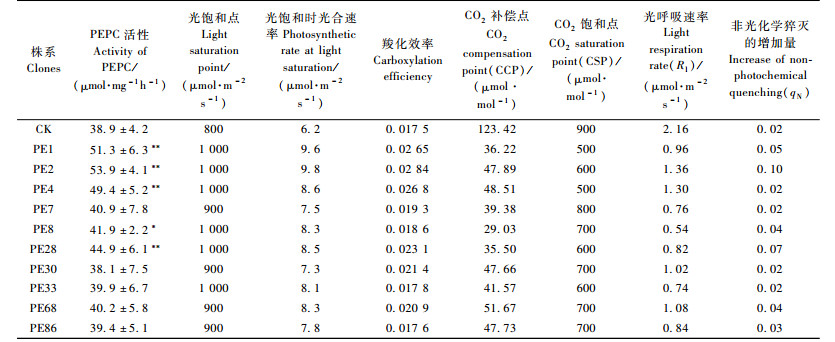
2 结果与分析 2.1 转PEPC基因南林895杨和对照叶片内光合相关酶活性比较在350 μmol·m-2s-1光强、21%O2、350 μmol·mol-1CO2,20 ℃条件下,测定结果显示:转PEPC基因南林895杨植株叶片中磷酸烯醇式丙酮酸羧化酶(PEPC)活性大多高于对照南林895杨,最高增加达39%(表 1)。除少数株系外,丙酮酸磷酸双激酶(PPDK)、NADP-苹果酸酶(NADP-ME)、NADP-苹果酸脱氢酶(NADP-MDH)活性也高于对照南林895杨,最高分别增加了25.7%,19.4%,19.1%。而C3光合酶RuBPc的活性与对照较为相近(表 1)。表 1中用PEPC/RuBPc比值描述测试杨树在C3和C4光合途径中的协调作用情况,与对照相比,转PEPC基因杨树最高上升32.3%。分析结果表明:转PEPC基因杨树中C4光合途径的关键酶磷酸烯醇式丙酮酸羧化酶(PEPC)活性增加较明显,特别是在PE1,PE2,PE4,PE28四个转基因株系中,PEPC酶活性分别比对照提高31.9%,38.6%,27.0%,15.4%,且均达到极显著差异水平(P < 0.01)。而C3光合酶RuBPc的活性与对照比较变化较小。
|
|

对转PEPC基因南林895杨和对照净光合速率进行测定结果表明:转PEPC基因杨树的净光合速率(Pn)日变化基本呈单峰曲线,而对照杨树的净光合速率(Pn)日变化呈双峰曲线,这与许大全(1990)提出的光合作用“午睡”现象类似。转PEPC基因南林895杨和对照杨树在早晚的净光合速率(Pn)基本类似。转PEPC基因杨树日变化呈单峰曲线,高峰出现在12:00左右,一天中最高净光合速率(Pn)达到14.3 μmol·m-2s-1,持续2 h左右开始下降。但对照杨树净光合速率日变化呈双峰曲线,高峰分别出现在11:00和14:00左右,达到12.7和12.8 μmol·m-2s-1,双峰曲线的低谷分别出现在12:00—13:00之间,持续1 h左右开始回升(表 2)。这表明转PEPC基因杨树能够利用中午的强光进行光合作用,而对照杨树在中午出现光抑制现象。
|
|
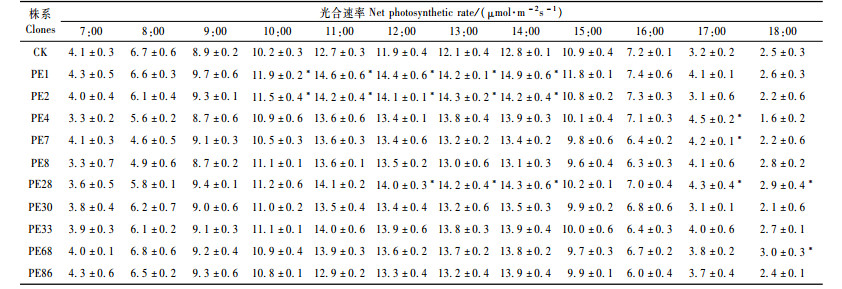
对转PEPC基因南林895杨和对照蒸腾速率进行测定结果表明:转基因杨树与对照的蒸腾速率(Tr)日变化基本均呈单峰曲线(表 3)。两者在上午时匀速上升,高峰均出现在14:00左右,转基因株系PE4 1天中最高蒸腾速率(Tr)达到6.8 mol·m-2s-1,对照植株1天中最高蒸腾速率(Tr)为5.8 mol·m-2s-1,两者之后迅速下降。表明大多数转PEPC基因杨树在中午的强光作用下蒸腾作用加强,而对照植株的蒸腾作用在中午有所抑制。
|
|
对转PEPC基因南林895杨和对照胞间CO2浓度进行测定结果显示:转基因杨树与对照的胞间CO2浓度日变化曲线基本一致(表 4)。两者在早晚时,胞间CO2浓度较高,高峰均出现在7:00与晚上18:00,峰值均在340 μmol·mol-1左右,在上午7:00后两者均有所下降。转PEPC基因南林895杨在中午时保持较高水平,而对照下降较快。上述分析表明两者的胞间CO2浓度日变化基本相同,并无明显差异。
|
|

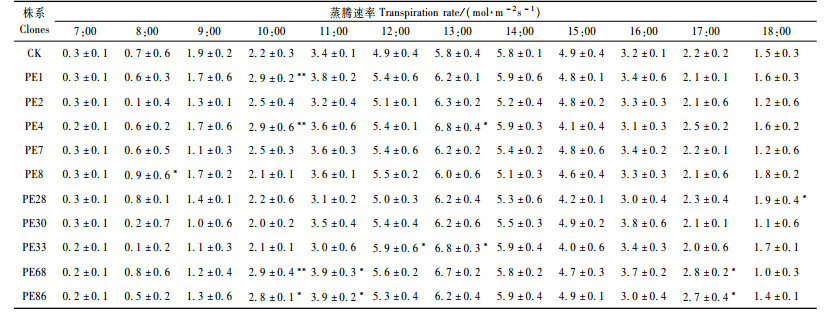
通常利用净光合速率(Pn)与光合有效辐射(PAR)的比值来表示光能利用效率(马成仓等,2004)。转基因杨树叶片光能利用效率日变化呈现双峰曲线,以上午9:00和下午15:00左右为最高,为2.9和2.8 mmol CO2·mol-1 photons,在中午时的光能利用效率比较低,最低值为1.0 mmol CO2·mol-1 photons。对照杨树叶片光能利用效率也是呈现双峰曲线,以上午9:00和下午15:00左右为最高,为2.4和2.5 mmol CO2·mol-1 photons,2个数值均比转基因杨树的略低;在中午时,对照的光能利用效率比转基因杨树略低,最低值为0.8 mmol CO2·mol-1 photons(表 5),显示转PEPC基因杨树利用强光的能力较强,在强光下依然保持高于对照的光能利用效率。
|
|
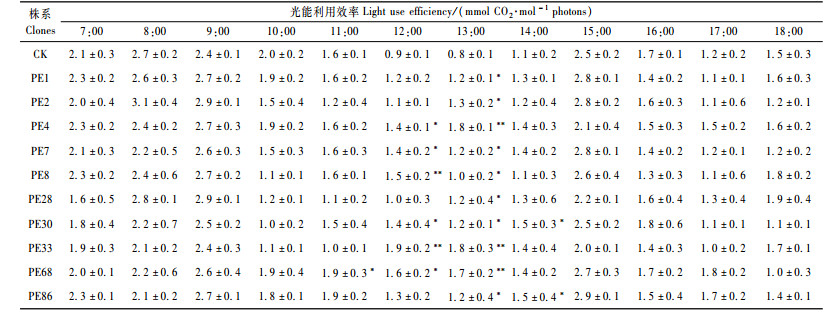
利用净光合速率(Pn)与蒸腾速率(Tr)的比值来表示瞬间水分利用效率(郑淑霞等,2006)。转基因杨树叶片的水分利用效率日变化以7:00最高,最高值为12.9 μmol CO2·mmol-1 H2O,然后逐渐下降;最低值出现在18:00,为1.0 μmol CO2·mmol-1H2O。对照的水分利用效率日变化与转基因杨树基本一致,最高值同样出现在7:00,为12.1 μmol CO2·mmol-1H2O,然后逐渐下降;最低值出现在18:00,为1.5 μmol CO2·mmol-1H2O(表 6)。水分利用效率的变化与蒸腾速率的变化趋势一致,表明水分利用效率在很大程度上决定于蒸腾速率,而两者的差异不明显。
|
|

光照强度对植物的光合作用影响显著。光合作用对光的响应测定结果表明(表 7),当光照强度在0~400 μmol·m-2s-1左右时,转PEPC基因杨树与对照的净光合速率几乎都呈线性增长。随着光照强度的不断增加,转PEPC基因杨树净光合速率的增长速度缓慢。当光照强度达到光饱和点以后,净光合速率几乎不增长。
|
|

根据叶片光响应变化,在光照强度0~400 μmol·m-2s-1范围内,净光合速率与光照强度呈线性关系,建立直线回归方程,计算得出每个材料的光饱和点、光补偿点、表观量子效率、暗呼吸速率以及光饱和时的光合速率(表 8)。
|
|
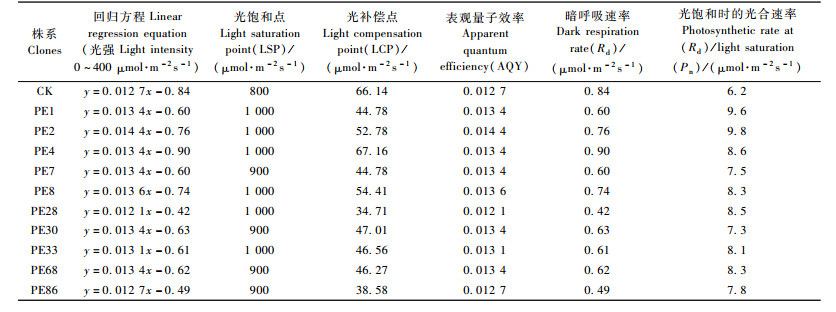
测定结果分析表明:在低光强条件下转PEPC基因杨树与对照的光合速率相近, 但随着光强的增加,光合速率在转PEPC基因杨树与对照间开始出现差异,转PEPC基因杨树的光饱和点比对照增加约10%~20%。转PEPC基因杨树在光饱和点时的光合速率比对照增高17.7%~58.1%。高光强可能诱导转基因杨树PEPC的高表达,加强C4光合产物的代谢能力,使光合速率得到提高。此外,转PEPC基因杨树的光补偿点除4号株系外,其他转基因株系均略低于对照杨树, 表明转PEPC基因杨树利用弱光的能力也较强。
2.6 转PEPC基因杨树与对照的光合作用对CO2浓度的响应比较CO2是光合作用的底物,CO2浓度高低直接影响植物的光合作用。CO2响应测定分析结果表明: CO2的浓度在0~400 μmol·mol-1范围内,转PEPC基因杨树与对照的净光合速率几乎呈线性增长(表 9)。但随着CO2浓度的不断增加,净光合速率的增长速度缓慢并最终趋于饱和。
|
|
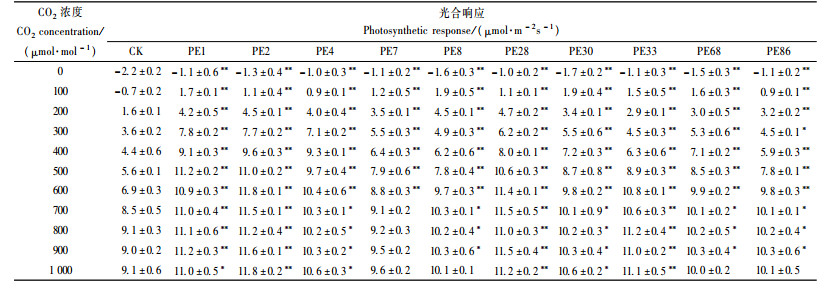
根据CO2浓度响应的变化,CO2浓度在0~400 μmol·m-2s-1范围内,净光合速率与CO2浓度呈线性关系,建立直线回归方程,计算得出每个试验材料的CO2饱和点、CO2补偿点、羧化效率、光呼吸速率、CO2饱和时的光合速率(表 10)。
|
|

分析结果显示:转PEPC基因杨树的CO2饱和点比对照低22.2%~44.4%,但在饱和点时的光合速率却比对照高,其中最高为2号株系,比对照增高29.7%(表 10)。转PEPC基因杨树的CO2饱和点均比对照低,在略高于大气CO2浓度下就达到饱和,而对照杨树CO2饱和点不明显,光合速率在较高CO2浓度下还会随浓度上升而提高。转PEPC基因杨树的Km低,CO2补偿点比对照低58.1%~76.5%,对CO2亲和力高,可能存在CO2浓缩机制,这些也是转PEPC基因杨树CO2饱和点低的原因。在低CO2浓度下转PEPC基因杨树光合速率的增加比对照快,CO2的利用效率较高。此外,转PEPC基因杨树的光呼吸比对照低37.0%~75.0%,光呼吸消耗较少,有利于光合作用有机物的积累,进而提高其生物量。分析结果显示:转PEPC基因杨树羧化效率比对照高,其中PE2株系最高,比对照增高62.3%。转PEPC基因杨树羧化效率较高,利用CO2的能力较强。
转PEPC基因杨树的CO2饱和点时的净光合速率(Pn),最高较对照提高了29.7%;光饱和时的净光合速率(Pn)较对照提高了17.7%~58.1%。分析结果表明:转PEPC基因杨树可能具备C4光合循环,也体现出一些C4植物的光合循环特性(表 11)。
|
|
本研究有关C4光合途径相关酶活性分析,与对照相比,转PEPC基因杨树表现有较高的PEPC酶活性。而转PEPC基因植株与对照植株在与C4光合途径其他相关的酶如丙酮酸磷酸双激酶、NADP-苹果酸酶、NADP-苹果酸脱氢酶(NADP-MDH)等活性方面相差不明显。光合作用参数分析结果表明:转PEPC基因杨树能够较好地利用中午的强光进行光合作用,光饱和点较高,且在光饱和点时的光合速率也较高,而对照植株在中午光饱和点时出现较为明显的光抑制现象;转PEPC基因杨树在中午的强光作用下蒸腾作用有所加强,叶片呼吸与散热能力有一定增强。而对照植株的蒸腾作用在中午有所抑制;转PEPC基因杨树在强光作用下,胞间CO2浓度依然维持较高水平,利用CO2的能力较对照植株高;与对照相比,转PEPC基因杨树利用弱光的能力较强,在一定程度上能够有效地利用弱光;转PEPC基因杨树羧化效率较高,利用CO2的能力较强,对CO2亲和力高,可能有浓缩CO2机制。转基因杨树PEPC活力与光饱和点时的光合速率极显著相关,PEPC活力与CO2饱和点时光合速率显著相关,这进一步证明了转PEPC基因杨树光合速率的提高与PEPC基因的导入有关。朱素琴等(2004)对转PEPC基因水稻(Oryza sativa)和野生型水稻叶内光合相关酶活性进行分析,PEPC基因水稻中除磷酸烯醇式丙酮酸羧化酶活性明显高于野生型水稻外,丙酮酸磷酸双激酶(PPDK)、NADP-苹果酸酶、NADP-苹果酸脱氢酶、碳酸酐酶以及C3光合酶RuBPc的活性均与野生型水稻相近。本研究结果与该研究结果类似。
3.2 讨论在植物长期进化过程中,C4植物形成与其光合循环相匹配的Kranz解剖结构。但一般认为C4植物光合作用的关键并不在于它的Kranz解剖结构,而主要决定于控制其C4途径的酶系统。近年来,据报导在不存在Kranz解剖结构的水生植物Hydrilla verticilata和Egeria densa中也发现有C4光合途径,预示着通过C4光合酶基因向C3植物的转移提高C3植物光合作用的可行性(Teng et al., 2001;Magnin et al., 1997;Li et al., 2005)。从Ku等(1999)利用农杆菌介导法成功地将玉米光合作用C4途径的关键酶基因PEPC导入水稻进行高光效分子育种以来,超表达的转PEPC基因水稻具有C4植物的高光效特性,并且在农艺性状及其最终产量表现优于对照(焦德茂等,2001;李霞等,2001;Jiao et al., 2002;Wang et al., 2002;Raines,2003;Wang et al., 2004;Raines et al., 2006;Price et al., 2008)。这为今后利用转基因技术开展杨树高光效分子育种研究提供重要的试验依据。
本研究利用转基因技术,将C4植物光合途径关键酶基因PEPC导入C3植物杨树中,初步分析结果显示:转PEPC基因杨树具有C4植物的高光效特性,改善了杨树C3光合作用循环,说明通过增强C4光合特性改良C3植物杨树光合作用以提高产量的可能,为超高产杨树育种开辟了一条新途径。但C4光合作用途径是极其复杂的生理生化过程,涉及许多基因的协同调控(Christine, 2011;程建峰等,2010;Murchie et al., 2009;Parry et al., 2007)。本研究仅利用单个C4光合作用关键基因进行杨树的遗传转化研究,结果也表明转PEPC基因杨树中与C4光合作用途径相关的酶只有磷酸烯醇式丙酮酸羧化酶(PEPC)获得较明显增高,而其他C4光合作用途径相关的酶没有显著变化。因此,光合效率的提高还需要在一系列C4光合途径相关酶系统研究的基础上,进行多个基因的分子聚合育种研究。
| [] | 程建峰, 沈允钢. 2010. 作物高光效之管见. 作物学报, 36(8): 1235–1247. |
| [] | 焦德茂, 李霞, 黄雪清, 等. 2001. 转PEPC基因水稻的光合CO2同化和叶绿素荧光特性. 科学通报, 46(5): 414–418. |
| [] | 李霞, 焦德茂, 戴传超, 等. 2001. 转PEPC基因的杂交水稻的光合生理特性. 作物学报, 21(2): 137–142. |
| [] | 马成仓, 高玉葆, 王金龙, 等. 2004. 内蒙古高原甘蒙锦鸡儿光合作用和水分代谢的生态适应性研究. 植物生态学报, 28(3): 306–311. |
| [] | 许大全. 1990. 光合作用"午睡"现象的生态、生理与生化. 植物生理通讯(6): 5–10. |
| [] | 郑淑霞, 上官周平. 2006. 8种阔叶树种叶片气体交换特征和叶绿素荧光特征比较. 植物生态学报, 26(4): 1080–1087. |
| [] | 朱素琴, 季本华, 焦德茂. 2004. 外源C4二羧酸对转玉米PEPC基因水稻C4光合途径的促进作用. 中国水稻科学, 18(4): 326–332. |
| [] | Bar-Even A, Noor E, Lewis N E, et al. 2010. Design and analysis of synthetic carbon fixation pathways. Proc Natl Acad Sci USA, 107(19): 8889–8894. DOI:10.1073/pnas.0907176107 |
| [] | Berg I A, Kockelkorn D, Ramos-Vera W H, et al. 2010. Autotrophic carbon fixation in archaea. Nat Rev Microbiol, 8(2): 447–460. |
| [] | Chen J Z, Chen D L, Shi J N. 1981. Comparison of some characteristics of NADP malic enzyme from sorghum and wheat leaves. Acta Phytophysiol Sin, 7(4): 345–350. |
| [] | Christine A R. 2011. Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: current and future strategies. Plant Physiology, 155(1): 36–42. DOI:10.1104/pp.110.168559 |
| [] | Edgerton M D. 2009. Increasing crop productivity to meet global needs for feed, food, and fuel. Plant Physiol, 149(1): 7–13. DOI:10.1104/pp.108.130195 |
| [] | Gonzalez D H, Iglesias A A, Andeo C S. 1984. On the regulation of phosphoenol pyruvate carboxylase activity from maize leaves by L-malate: effect of pH. J Plant Physiol, 116(2): 425–429. |
| [] | Jiao D M, Huang X Q, Li X, et al. 2002. Photosynthetic characteristics and tolerance to photo-oxidation of transgenic rice expressing C4 photosynthesis enzymes. Photosynthesis Res, 72(1): 85–93. DOI:10.1023/A:1016062117373 |
| [] | Ku M S B, Kano-Murakami Y, Matsuoka M. 1996. Evolution and expression of C4 photosynthesis genes. Plant Physiol, 111(5): 949–957. |
| [] | Ku M S B, Sakae A, Mika N, et al. 1999. High level expression of maize phosphoenol pyruvate carboxylase in transgenic rice plants. Nat Biotechnoly, 17(1): 76–80. DOI:10.1038/5256 |
| [] | Kung S D, Chollet R, Marsho T V. 1980. Crystallization and assay procedures of tobacco ribulose-1, 5-bisphosphate carboxlase-oxygenase//Pietro A S. Method in Enzymology. New York, Academic Press Inc: 326-335. |
| [] | Li B, Chen D L, Shi J N. 1987. Purification and molecular properties of NADP dependent malate dehydrogenase from sorghum leaves. Acta Phytophysiol Sin, 13(2): 113–121. |
| [] | Li X, Jiao D M. 2005. Transgenic rice overexpressing C4 photosynthetic genes and their application in breeding. Mol Plant Breed, 3(4): 550–556. |
| [] | Magnin N C, Cooley B A, Reiskind J B, et al. 1997. Regulation and localization of key enzymes during the induction of Kranzless, C4 type photosynthesis in Hydrilla verticillata. Plant Physiol, 115(4): 1681–1689. DOI:10.1104/pp.115.4.1681 |
| [] | Moore P. 1982. Evolution of photosynthetic pathways in flowering plants. Nature, 295(5893): 647–648. |
| [] | Murchie E H, Pinto M, Horton P. 2009. Agriculture and the new challenges for photosynthesis research. New Phytol, 181(3): 532–552. DOI:10.1111/j.1469-8137.2008.02705.x |
| [] | Olcer H, Lloyd J C, Raines C A. 2001. Photosynthetic capacity is differentially affected by reductions in sedoheptulose-1, 7-bisphosphatase activity during leaf development in transgenic tobacco plants. Plant Physiol, 125(2): 982–989. DOI:10.1104/pp.125.2.982 |
| [] | Parry M A J, Madgwick P J, Carvalho J F C. 2007. Prospects for increasing photosynthesis by overcoming the limitations of Rubisco. J Agric Sci, 145(1): 31–43. DOI:10.1017/S0021859606006666 |
| [] | Portis A R Jr, Parry M A J. 2007. Discoveries in Rubisco (ribulose 1, 5-bisphosphate carboxylase/oxygenase): a historical perspective. Photosynth Res, 94(1): 121–143. DOI:10.1007/s11120-007-9225-6 |
| [] | Price G D, Badger M R, Woodger F J, et al. 2008. Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J Exp Bot, 59(7): 1441–1461. DOI:10.1093/jxb/erm112 |
| [] | Raines C A. 2003. The Calvin cycle revisited. Photosynth Res, 75(1): 1–10. DOI:10.1023/A:1022421515027 |
| [] | Raines C A, Paul M J. 2006. Products of leaf primary carbon metabolism modulate the developmental programme determining plant morphology. J Exp Bot, 57(9): 1857–1862. DOI:10.1093/jxb/erl011 |
| [] | Sage R F. 2002. Variation in the kcat of Rubisco in C3 and C4 plants and some implications for photosynthetic performance at high and low temperature. J Exp Bot, 53(369): 609–620. DOI:10.1093/jexbot/53.369.609 |
| [] | Teng S, Qian Q, Huang D N. 2001. Advance in molecular biology and genetic engineering of C4 photosynthesis pathway. J Agric Biotechnol, 9(2): 198–201. |
| [] | Wei J C, Wang R L, Cheng G Y. 1994. Studies on the kinetic properties of ribulose-1, 5-biphosphate carboxylase from F1 hybrid rice. Acta Phytophysiol Sin, 20(1): 55–60. |
| [] | Wang D Z, Jiao D M, Wu S, et al. 2002. Breeding for parents of hybrid rice with maize PEPC gene. Sci Agric Sin, 35(10): 1165–1170. |
| [] | Wang D Z, Chi W, Wang S H, et al. 2004. Characteristics of transgenic rice over-expressing maize photosynthetic enzymes for breeding two line hybrid rice. Acta Agron Sin, 30(3): 248–252. |