 2012, Vol. 48
2012, Vol. 48文章信息
- 邵方丽, 余新晓, 吴海龙, 黄枝英
- Shao Fangli, Yu Xinxiao, Wu Hailong, Huang Zhiying
- 冀北土石山区天然次生林山杨、白桦种群的空间分布格局
- Spatial Distribution Pattern of Populus davidiana and Betula platyphylla Populations in a Typical Natural Secondary Forest in Rocky Mountain Area of Northern Hebei
- 林业科学, 2012, 48(6): 12-17.
- Scientia Silvae Sinicae, 2012, 48(6): 12-17.
-
文章历史
- 收稿日期:2011-04-28
- 修回日期:2012-04-20
-
作者相关文章



目前,国内对天然次生林的研究主要集中于林隙特征(李雪芬等,2008;符利勇等,2010;2011)、林下土壤(林丽莎等,2009;侯琳等,2008)及林下幼苗更新(宋新章等,2008)等方面,对于空间分布及空间关联的研究较少(闫海冰等,2010)。空间分布格局是种内/种间相互关系及环境条件综合作用的结果,空间格局分析是研究种群结构特征、种群相互作用以及种群与环境间关系的重要方法(王巍等,1999)。种群在不同尺度下呈现出不同的空间格局(王本洋等,2005),Ripley's K函数可以进行多尺度空间格局以及空间关联分析,能更好地反映种群空间特征,被广泛应用于种群的空间格局分析中(张金屯等,1998;2004)。本研究分析冀北土石山区天然次生阔叶林中山杨、白桦种群的径级结构、空间分布格局及种内/间不同生长阶段个体间的空间关联,以期为天然次生林退化生态系统的恢复与重建提供依据。
1 研究区概况研究区位于河北省木兰围场国有林场管理局所辖的北沟林场(116°32′—117 °14′E,41°35′—42°40′N)。北沟林场位于围场满族蒙古族自治县境内,地处浑善达克沙地南缘,滦河上游地区,属阴山、大兴安岭、燕山余脉的汇接地带,海拔750~1 998 m,属半干旱向半湿润过渡、寒温带向中温带过渡的大陆季风型山地气候,全年无霜期67~128天,年平均气温-1.4~4.7 ℃,极端最高气温38.9 ℃,极端最低气温-42.9 ℃,年均降水量380~560 mm,主要集中在7—9月。森林覆被率85.6%,活立木蓄积20.9万m3。北沟林场有天然林1 485.4 hm2,主要树种为山杨(Populus davidiana)、白桦(Betula platyphylla)、华北落叶松(Larix principis-rupprechtii)和云杉(Picea asperata)等;人工林1 180.5 hm2,主要树种为油松(Pinus tabulaeformis)和樟子松(Pinus sylvestris var. mongolica)等。本研究中的天然杨桦次生林是在暖温带落叶阔叶林和温带针叶林遭受火灾干扰后,在火烧迹地上自然恢复而形成的,该次生林以山杨、白桦为先锋树种,并伴生有色木槭(Acer mono)和华北落叶松等,林下生长有大量的毛榛(Corylus mandshurica)灌丛。该群落在20世纪60—70年代经历了对大中径木的较大强度择伐,以后处于自然恢复状态。
2 研究方法 2.1 样地设置与调查2010年7—9月设置1块200 m×200 m的天然杨桦次生林大样地,大样地位于阳坡,平均坡度20°,海拔1 330~1 440 m。用TOPCON全站仪设定大样地的坐标基轴,再用全站仪将整个样地划分为100块20 m×20 m小样地。以小样地为基本单元,对大样地内所有DBH≥1 cm的乔木进行调查,调查内容包括物种、胸径、树高、冠幅(东西冠幅长、南北冠幅长)、枝下高及其位置坐标。
2.2 数据分析径级结构是指乔木在不同胸径(DBH)级上个体数的分布情况。本研究径级结构划分方法如下:DBH≤2 cm为径阶1,2 cm<DBH≤4 cm为径阶2,4 cm<DBH≤6 cm为径阶3,6 cm<DBH≤8 cm为径阶4,8 cm<DBH≤10 cm为径阶5,以此类推,此后均以2 cm为1个径阶来进行林分径级结构划分。
空间格局分析采用Ripley's K(t)函数点格局分析方法。该函数用来描述不同空间尺度下的种群分布格局。其公式为:
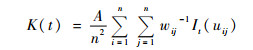
|
(1) |
式中:A为样地面积;n为植物个体总数;uij为2个点i和j之间的距离;wij是以i为圆心,uij为半径的圆周长落在样地内的长度与圆周长的比例;t为距离尺度;It(uij)为指示函数,当uij≤t时,It(uij)=1,当uij>t时,It(uij)=0。为保持方差稳定,Besag等提出用L(t)取代K(t)(Besag,1977),L(t)计算公式如下:
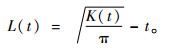
|
(2) |
当L(t)=0时,为随机分布;当L(t)>0时,为聚集分布;当L(t)<0时,为均匀分布。采用Monte-Carlo模拟99%置信区间,进行结果偏离随机状态的显著性检验。L(t)值位于置信区间之上,种群呈聚集分布;L(t)值位于置信区间之下,种群呈均匀分布;L(t)值位于置信区间之内,种群呈随机分布。
当种群表现为聚集分布时,把偏离随机置信区间的最大值定义为聚集强度,对应于聚集强度的尺度定义为聚集尺度,以聚集尺度为半径的圆面积定义为最大聚集规模(Sotirios et al., 2005)。
种内及种间空间关系的分析采用Ripley's K12(t)函数确定。其公式为

|
(3) |
式中:n1,n2分别为2个类型组的个体数。为保持方差稳定,用L12(t)取代K12(t)。L12(t)计算公式如下:

|
(4) |
当L12(t)=0时,2个变量相互独立;L12(t)>0时,2个变量空间正关联;当L12(t)<0时,2个变量空间负关联。采用Monte-Carlo模拟99%置信区间,当L12(t)值位于置信区间之上时,2个变量显著正相关;L12(t)值位于置信区间之下时,2个变量显著负相关;L12(t)值位于置信区间之内,2个变量相互独立。
Ripley's K函数分析通过SPPA软件实现。研究尺度的最大值设为样地最短边的1/2(100 m),步长设为1 m。
3 结果与分析 3.1 乔木树种组成该天然次生林群落的主要乔木树种有山杨、白桦、华北落叶松、色木槭、花楸(Sorbus pohuashanensis)、蒙古栎(Quercus mongolica)、青杄(Picea wilsonii)和棘皮桦(Betula duburica)等,重要值从大到小排前5位的分别是山杨、白桦、华北落叶松、色木槭和花楸,山杨和白桦的重要值分别为33.98和27.70,其个体总株数分别占样地乔木总株数的41.6%和29.7%,胸高断面积分别占总断面积的38.3%和32.4%。
在相同的环境条件下,同一树种的径级和龄级对环境的反应规律具有一致性(Johnson,1997),本研究采用径级结构代替年龄结构来分析种群分布格局。根据胸径将树木划分为幼树(1 cm≤DBH≤5 cm)、小树(5 cm<DBH<25 cm)和大树(DBH≥25 cm)。调查共测得山杨1 301株,其中幼树42株,小树1 142株,大树117株;白桦927株,其中幼树171株,小树590株,大树166株。山杨、白桦个体在样地中的分布情况如图 1。

|
图 1 山杨、白桦的散点分布 Fig.1 Scatter distribution diagrams of P. davidiana and B. platyphylla |
从树种的径阶结构(图 2)来看,山杨、白桦种群径级分布范围较广,山杨的平均胸径为16.1 cm,最大胸径为47.4 cm;白桦的平均胸径为16.1 cm,最大胸径为47.5 cm。山杨的径级呈近似正态分布:在1~24径级范围内呈连续分布,1~9径级范围内随着径级的增大个体数逐渐增多,径级9个体数最多,之后随径级的增大个体数逐渐减少,17径级后的个体数大幅减少,多数径级上仅1个。白桦的径级呈多峰状分布,在径级2上出现第1个也是最大峰值,个体数为140,其他径级上的个体数均在100以下,在10~24径级上个体数随径级的增大逐渐减少,19径级后的个体数大幅减少,各径级上的个体数仅几个。

|
图 2 山杨(a)、白桦(b)的径级分布 Fig.2 DBH class distribution of P. davidiana (a) and B. platyphylla (b) |
径阶结构中,径级1~4的山杨、白桦的个体数分别占各自个体总数的12.9%和22.1%;径级5~13的中径木个体数分别占80.2%和62.5%;径级>13的大径木个体数分别占6.9%和15.4%。2个树种的小径木数量少,均以中径木为主,表明该群落优势种群缺乏可供更新的幼苗、幼树,种群呈现衰退趋势。林分大径木少,与该林区早期进行的大、中径木较大强度择伐所致。
3.3 种群在不同生长阶段上的空间分布Ripley's L(t)函数分析结果(图 3)表明:山杨全部个体在研究尺度上呈聚集分布,聚集强度为11.354 8,聚集规模为15 836.77 m2;山杨幼树和小树在本研究尺度上均呈聚集分布,聚集强度分别为31.145 3和11.926 2,聚集规模分别为11 309.73 m2和14 957.12 m2;大树在3~100 m尺度上为聚集分布,聚集强度为12.482 8,聚集规模为4 778.36 m2,在0~2 m尺度上呈随机分布;不同生长阶段的山杨,随尺度的增大聚集强度先增大后减小,趋于随机分布,但在本研究尺度上未过渡为随机分布。

|
图 3 山杨和白桦在不同生长阶段上的点格局分析 Fig.3 Point pattern analysis for individuals of Populus davidiana and Betula platyphylla of different growth stages 图中实线为L(t)的值,表示种群的分布情况;虚线为包迹线,表示所模拟的99%置信区间。 Solid line is the value of L(t), indicate the distribution of the population; dotted line is envelope line, indicate the simulated 99% confidence interval. |
由图 3还可知:白桦全部个体在研究尺度上均为聚集分布,聚集强度为9.856 2,聚集规模为8 494.87 m2;白桦幼树在所有尺度上均呈聚集分布,聚集强度为58.679 5,聚集规模为10 935.88 m2;小树在0~97 m尺度上呈聚集分布,最大聚集强度为7.417 0,聚集规模为7 238.23 m2,在其余尺度上呈随机分布;大树在5~98 m尺度上呈聚集分布,聚集强度为5.576 8,聚集规模为7 853.98 m2,在其余尺度上为随机分布。
3.4 种内不同生长阶段个体的空间关联Ripley's L12(t)双变量空间关联分析结果见表 1。山杨种群内的空间关联表现为:幼树与小树在0~1 m尺度上显著正关联,在其余尺度上相互独立;幼树与大树在0~2 m和90~100 m尺度上显著正关联,在3~76 m尺度上显著负关联,在77~89 m尺度上相互独立;小树与大树在0~90 m尺度上显著负关联,随尺度的增加负关联逐渐减弱,在91~100 m尺度上相互独立。白桦种群内的空间关联表现为:幼树与小树在0~35 m尺度上相互独立,在36~100 m尺度上显著正关联,在73 m尺度上正关联达到最大;幼树与大树在所有研究尺度上显著负关联,在34 m尺度上负相关达到最大;小树与大树在所有尺度上显著负关联。
|
|
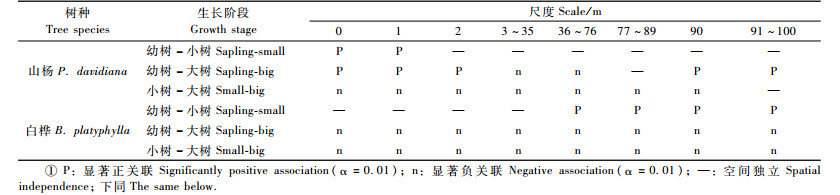
种间空间关联分析结果见表 2。山杨幼树与白桦小树在0~1 m尺度上相互独立,其余尺度上显著负关联,在87 m尺度上负关联最大;山杨幼树与白桦大树在19~96 m尺度上显著负关联,最大负关联不明显,其余尺度上相互独立;其余种对在本研究尺度上均为显著负关联。其中,山杨幼树与白桦幼树、山杨小树与白桦幼树在本研究尺度上未出现最大负关联,山杨小树与白桦小树在74 m尺度上负关联最大,山杨小树与白桦大树在54 m尺度负关联最大,山杨大树与白桦幼树在86 m尺度负关联最大,山杨大树与白桦小树在56 m尺度负关联最大,山杨大树与白桦大树未出现明显的最大负关联尺度。山杨与白桦的全部个体在本研究尺度上呈显著负关联,83 m尺度上的负关联达到最大;总体来看,在本研究尺度上未出现任何显著正关联,表明山杨与白桦各生长阶段的个体间存在着激烈的竞争。
|
|

研究表明,山杨、白桦的全部个体在本研究尺度上均呈聚集分布,种群的聚集分布有利于种群发挥群体效应,抵御外来种的入侵,形成适于自身生长的环境,从而维持种群的正常发展(韩路等,2007;张金屯等,2004)。种群空间格局与空间尺度紧密相关(He et al., 2000)。本研究中山杨、白桦种群的聚集强度随尺度增加而减弱,并逐渐转向随机分布,白桦大树随尺度增加出现随机分布的格局。种群空间格局随尺度的变化受多种因素制约,在较小尺度上,不同的空间格局可能是由植物繁殖特性、种子散布方式、群落种内或种间竞争等因素所致;在较大尺度上,不同的空间格局则可能是由生境(地形、水分、养分等)的异质性所决定(张春雨等,2008)。
种群内,山杨幼树与大树、白桦幼树与小树在部分尺度上为正关联,原因可能是同种间一个生长阶段的个体为另一个生长阶段个体的生存创造了条件,即它们对生境要求互补(蔡飞等,1997)。幼树与小、大树在部分尺度上的相互独立,可能与幼树的数量较少,且在样地中集中分布有关。种群内的多数种对在大部分尺度上呈负关联,则表明种群内不同生长阶段的个体间存在着激烈的竞争,群落结构不稳定。种群间,山杨、白桦的全部个体在本研究尺度上均为显著负关联;种群间不同生长阶段的个体在本研究尺度上无正关联,除山杨幼树-白桦小树、山杨幼树-白桦大树在某些尺度上为相互独立外,其余的种间关系均为显著负关联,体现出山杨、白桦种群间的相互排斥性。山杨、白桦作为该天然次生林的先锋树种,发生于群落演替初期,其种群内和种群间存在着极其激烈的竞争,导致群落整体结构十分不稳定。
种群的空间正/负关联性影响着群落的演替,在森林生态系统中,引起种群空间关系变化的原因较多,需进行更加深入的研究。本研究中山杨、白桦的空间分布格局及种内/种间空间关联性除受采伐干扰和树种繁殖特性的影响外,还受到其他因素影响(如林下大量生长的灌木等)。山杨、白桦种群与灌木的空间关联有待进一步研究。
| [] | 蔡飞, 宋永昌. 1997. 武夷山木荷种群结构和动态的研究. 植物生态学报, 21(2): 138–148. |
| [] | 符利勇, 何铮, 刘应安. 2010. 关帝山天然次生针叶林林隙大小模型研究. 南京林业大学学报:自然科学版, 4(5): 51–54. |
| [] | 符利勇, 唐守正, 刘应安. 2011. 关帝山天然次生针叶林林隙径高比. 生态学报, 31(5): 1260–1268. |
| [] | 韩路, 王海珍, 彭杰, 等. 2007. 塔里木河上游天然胡杨林种群空间分布格局与动态研究. 西北植物学报, 27(8): 1668–1673. |
| [] | 侯琳, 雷瑞德, 王得祥, 等. 2008. 秦岭火地塘天然次生油松林土壤有机碳的特征. 西北农林科技大学学报:自然科学版, 36(8): 156–159. |
| [] | 李雪芬, 韩有志, 张莉, 等. 2008. 关帝山天然次生针叶林的林隙特征. 浙江林学院学报, 25(1): 28–32. |
| [] | 林丽莎, 李向义. 2009. 韩士杰长白山天然次生白桦林土壤CO2释放通量研究. 干旱区地理, 32(1): 67–71. |
| [] | 宋新章, 张智婷, 肖文发, 等. 2008. 长白山杨桦次生林采伐林隙幼苗更新动态. 林业科学, 44(3): 13–20. DOI:10.11707/j.1001-7488.20080307 |
| [] | 王本洋, 余世孝. 2005. 种群分布格局的多尺度分析. 植物生态学报, 29(2): 235–241. DOI:10.17521/cjpe.2005.0030 |
| [] | 王巍, 刘灿然, 马克平, 等. 1999. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 41(4): 425–432. |
| [] | 闫海冰, 韩有志, 杨秀清, 等. 2010. 华北山地典型天然次生林群落的树种空间分布格局及其关联性. 生态学报, 30(9): 2311–2321. |
| [] | 张春雨, 赵秀海, 夏富才. 2008. 长白山次生林树种空间分布及环境解释. 林业科学, 44(8): 1–8. DOI:10.11707/j.1001-7488.20080801 |
| [] | 张金屯. 1998. 植物种群空间分布的点格局分析. 植物生态学报, 22(4): 344–349. |
| [] | 张金屯, 孟东平. 2004. 芦芽山华北落叶松林不同龄级立木的点格局分析. 生态学报, 24(1): 35–40. |
| [] | Besag J E. 1977. Comments on Ripley's paper. Journal of the Royal Statistical Society, 39(2): 193–195. |
| [] | He F, Duncan R P. 2000. Density-dependent effects on tree survival in an old-growth Douglas-fir forest. Journal of Ecology, 88(4): 676–688. DOI:10.1046/j.1365-2745.2000.00482.x |
| [] | Johnson J B. 1997. Stand structure and vegetation dynamics of a subalpine wooded fen in Rocky Mountain National Park, Colorado. Journal of Vegetation Science, 8(3): 337–342. DOI:10.2307/3237322 |
| [] | Sotirios K, George A B. 2005. Spatial relationships between tree species and gap characteristics in broadleaved deciduous woodland. Journal of Vegetation Science, 16(5): 587–596. DOI:10.1111/j.1654-1103.2005.tb02400.x |