 2012, Vol. 48
2012, Vol. 48文章信息
- 李会平, 黄大庄, 唐秀光
- Li Huiping, Huang Dazhuang, Tang Xiuguang
- 桑天牛幼虫感染球孢白僵菌后蛋白质和氨基酸含量的变化
- Changes in the Contents of Proteins and Amino Acids of Apriona germari Larvae Infected by Beauveria bassiana
- 林业科学, 2012, 48(5): 159-163.
- Scientia Silvae Sinicae, 2012, 48(5): 159-163.
-
文章历史
- 收稿日期:2010-09-12
- 修回日期:2011-05-07
-
作者相关文章


桑天牛现称桑粒肩天牛(Apriona germari),是多种林木、果树、花卉的重要蛀干害虫,也是我国危害最严重、最难防治的害虫之一。近年来,其猖獗危害已成为我国养蚕业、林业及果树种植业等经济发展的严重障碍。球孢白僵菌(Beauveria bassiana)是一类广谱性昆虫病原真菌,能侵染15目149科的700多种昆虫(St Leger,2001; Blake et al., 2002; Felipe et al., 2004)。近年来,利用白僵菌防治松毛虫(Dendrolimus spp.)、玉米螟(Pyrausta nubilalis)、棉铃虫(Helicoverpa armigera)、美国白蛾(Hyphantria cunea)、天牛等农林害虫的研究和应用均取得很大进展,在控制农林害虫的严重发生和减少环境污染方面,发挥着不可替代的作用(赫英伟,2010; 王滨,2003; 丁德贵等,2004; 张爱文等,1992; 孙鲁娟等,2001; 郭志红等,2001; 刘洪剑等,2007; Shmazu,1995; 徐均焕等,2003; 林庆源,2005; 刘洪剑等,2009; 王云滨,2007; 陆秀君等,2008)。李会平等(2006)从诱集自土壤中的9株白僵菌中初步筛选出了对桑天牛幼虫具有较高致病性的Bb00菌株,其田间防治效果良好。球孢白僵菌通过分生孢子的附着、萌发、穿透和在体腔中增殖,进行营养生长。此过程中,对感染昆虫最重要的生理作用是对血淋巴的影响。入侵到血腔里的真菌一般都能在血腔内依靠血淋巴成分进行增殖,使昆虫出现生理饥饿; 因此患病昆虫体内氨基酸含量会降低。但是菌株的入侵也必然会受到昆虫机体免疫反应的抵御,促使寄主血淋巴中蛋白质分解成氨基酸,以弥补由于菌株营养生长所造成的寄主体内氨基酸含量的减少,从而维持虫体的生长发育,使幼虫不会由于“生理饥饿”而立即死亡。同时由于昆虫的血淋巴含有来自各种组织和血细胞等合成和分泌的蛋白质、氨基酸和酶类(傅丽君,2000; 牛宇,2005),因此随着白僵菌在体内的增殖和扩展,罹病昆虫体内的蛋白质和氨基酸含量必然会出现一系列变化。薛皎亮等(2006)采用分光光度法测定了油松毛虫(D. tabulaeformis)3~5龄幼虫被白僵菌感染后体内蛋白质含量的变化,结果表明,白僵菌感染后1~8天,虫体内蛋白质含量持续下降,而未感染的则变化不大。付丽君等(2001)通过高效液相色谱(HPLC)对感染球孢白僵菌后小菜蛾(Plutella xylostella)血淋巴中游离氨基酸含量进行了测定,发现小菜蛾幼虫体内具有菌株营养生长所必需的氨基酸,染菌后小菜蛾幼虫体内氨基酸含量明显低于对照组,且各处理组间各种游离氨基酸在量上存在一定差异。李庚祥等(2009)的研究表明球孢白僵菌及粉拟青霉(Paecilomyces farinosus)对松墨天牛(Monochamus alternatus)幼虫的毒力有一定的差异,侵染三龄幼虫后,可造成虫体血淋巴中蛋白质含量降低,且球孢白僵菌侵染后血淋巴中总蛋白含量明显低于经粉拟青霉接种处理感染后的虫体。李会平(2007)和王达等(2009)通过扫描电镜并结合石蜡切片观察了白僵菌对桑天牛幼虫的侵染致病过程,明确了桑天牛幼虫血淋巴对球孢白僵菌的防御反应和免疫作用。但对于白僵菌侵染导致的昆虫免疫机制发挥作用这一过程中蛋白质和氨基酸的变化还鲜见报道,因此,本研究以感染白僵菌后的桑天牛幼虫为研究对象,结合白僵菌对桑天牛幼虫的侵染致病过程,综合分析了蛋白质和氨基酸含量变化与白僵菌侵染致病的关系,以期为明确白僵菌与桑天牛之间的互作关系,研究白僵菌的侵染和致病机制提供理论依据。
1 材料与方法 1.1 供试昆虫桑天牛幼虫:自河北省保定市清西陵捕捉桑天牛成虫利用1~2年生幼嫩桑枝进行室内饲养。产卵后10天左右,剖取已孵化的桑天牛幼虫,人工接种到毛白杨(Populus tomentosa)枝条中,以见蛀屑和虫粪排出确认成活,于接种后30天取幼虫用于以下测定。
1.2 供试菌株及孢子悬浮液制备球孢白僵菌Bb00菌株为诱集自土壤中对桑天牛幼虫具有较高致病性的菌株,由河北农业大学林学院森林保护教研室提供。利用扫描电镜和石蜡切片观察该菌的致病过程如下:白僵菌处理桑天牛幼虫后16 h开始萌发并出现芽状突起,随后产生芽管; 20~24 h孢子即可穿透体壁进入血腔。菌丝进入血腔后开始大量繁殖,并战胜昆虫的免疫系统,逐渐侵染体腔内的组织与器官(李会平,2007; 王晓红,2008)。
在察氏培养基(许文耀,2006)中培养的白僵菌完全产孢后,刮下分生孢子,用含0.1%吐温-80的无菌水润湿,磁力搅拌器搅拌均匀,经血球计数板计数后,配成1×108个·mL-1的孢子悬浮液。
1.3 蛋白质含量测定及电泳分析将30天龄桑天牛幼虫浸入孢子悬浮液中30 s后,接入毛白杨枝条中,分别于接种后1,2,3,4,5,6天取出受侵染幼虫,用蒸馏水冲洗3~4遍,滤纸吸干后在虫体稍前部用消毒的细昆虫针扎小口,当血淋巴从伤口溢出成小水珠状,用微量进样器吸取收集血淋巴于一干净离心管中(提前冷冻且加入少许苯基硫脲),然后在4 ℃,12 000 r·min-1,离心15 min后除去血细胞备用。蛋白质含量测定及电泳分析各重复3次。
蛋白质含量采用考马斯亮蓝G250法测定,用牛血清蛋白制作标准曲线(西北农业大学,1986)。
蛋白质组分分析采用Tris-甘氨酸电极缓冲系统,以垂直平板电泳槽(BIO-RAD公司)进行电泳。分离胶和浓缩胶浓度分别为7.5%和3%,pH值分别为8.9和6.7。每样品加样量为30 μL(以20%蔗糖溶液1:1与血淋巴上清液混合制成电泳样品液),电泳初始电压为80 V,待指示剂进入分离胶后将电压调至120 V,溴芬蓝线距分离胶底部1㎝时,停止电泳。
电泳完毕后,用考马斯亮兰G250染色2 h,10%冰乙酸脱色至背景白色透明。计算各蛋白质谱带迁移率。
迁移率(Rf)=谱带迁移距离/溴酚蓝迁移距离。
1.4 白僵菌侵染对桑天牛幼虫血淋巴氨基酸含量的影响同上方法收集血淋巴液50 μL,迅速加入100 μL3%的磺基水杨酸溶液(以上处理均在冰浴下进行)。4 ℃,12 000 r·min-1离心15 min,取上清液保存于-20 ℃冰箱中备用。
仪器:美国Waters2690高效液相色谱仪。
色谱条件:色谱柱:EliteAAK氨基酸专用柱4.6 mm×250 mm。
流动相:0.05 moL·L-1醋酸钠缓冲液,乙腈:水=1:1; 0~37 ℃梯度洗脱。
柱温27 ℃,紫外360 nm检测,运行时间45 min,进样量10 μL。
2 结果与分析 2.1 白僵菌侵染对桑天牛幼虫血淋巴蛋白质含量的影响白僵菌侵染对桑天牛幼虫血淋巴中蛋白质含量的影响见图 1。

|
图 1 桑天牛幼虫感染白僵菌后血淋巴蛋白质含量的变化 Fig.1 Changes of the content of protein in haemolym of A. germari larvae infected with B. bassiana |
从图 1可以看出:对照组桑天牛幼虫血淋巴中蛋白质含量变化不大; 而处理组桑天牛幼虫第1天蛋白质含量与对照组相比无显著差异。这主要由于白僵菌喷于虫体后,大量的孢子萌发、侵入需在24 h后才能完成,而少量早期侵入昆虫体内的白僵菌,其对营养的需求主要取自血液中的游离氨基酸。自第2天起,其蛋白质含量逐渐升高并高于对照组,至第3天达最高值,此时蛋白质含量为对照组的2.63倍; 之后又逐渐下降,至第6天时仅为对照组的1.44倍。
2.2 白僵菌侵染对桑天牛幼虫血淋巴蛋白质组分的影响感染和对照桑天牛幼虫血淋巴蛋白图谱见图 2。

|
图 2 桑天牛幼虫感染白僵菌后1~6天血淋巴蛋白电泳图谱 Fig.2 Polyacrylamide gel electrophoretic patterns of haemolymph proteins of A.germari larvae on 1st to 6th day after infection |
由图 2可以看出:白僵菌侵染后,桑天牛幼虫血淋巴蛋白谱带数量比对照有所增加,这与前面蛋白质含量测定的结果是一致的。其中,自侵染第1天起,出现了相对迁移率为0.047的b带,其颜色逐渐加深,一直到第5天该带又逐渐消失。自第2天起,相对迁移率为0.66的a带逐渐加宽,到第5天该带逐渐恢复到对照和第1天时的水平。相对迁移率为0.78的c带自第5天起颜色变浅。而在第5天和第6天分别新增了前期没有的d带,其相对迁移率为0.81。
2.3 白僵菌侵染对桑天牛幼虫血淋巴氨基酸含量的影响白僵菌侵染后桑天牛幼虫血淋巴中游离氨基酸含量的变化见表 1。
|
|
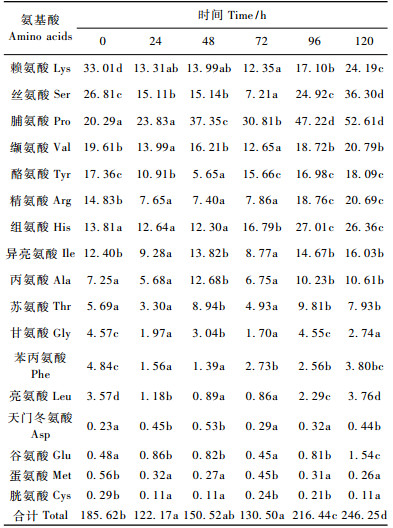
由表 1可知:桑天牛幼虫血淋巴中共检测出17种游离氨基酸,其中主要氨基酸有赖氨酸、丝氨酸、脯氨酸、缬氨酸、酪氨酸、精氨酸、组氨酸、异亮氨酸; 而天门冬氨酸、谷氨酸、蛋氨酸和胱氨酸总含量最低,为稀有氨基酸。白僵菌侵染未影响幼虫血淋巴中游离氨基酸的种类,但氨基酸含量发生了一定变化。
从氨基酸总量上看,感染24 h时,桑天牛幼虫血淋巴中的游离氨基酸总量明显下降,这主要是由于24 h时少量白僵菌侵入虫体吸收血液中游离氨基酸,而此时寄主的免疫调节还未被完全启动; 感染24~72 h时,游离氨基酸的总量变化不大,可能是寄主免疫反应参与的结果; 感染96 h和120 h时桑天牛幼虫血淋巴中游离氨基酸的总量急剧上升,分别为72 h时的1.69和1.87倍。分析其原因,主要与桑天牛幼虫自身免疫调节功能的逐步丧失以及组织被破坏、解离、蛋白质解体有关。
其次,不同种类氨基酸表现出不同的变化趋势。主要氨基酸中的赖氨酸、酪氨酸、精氨酸、丝氨酸和氨基酸总量的变化趋势一样,是影响氨基酸总量变化的主要氨基酸种类,而脯氨氨酸和组氨酸含量逐渐上升。其他氨基酸则没有明显规律,呈波动性变化。这说明白僵菌侵染桑天牛幼虫的不同阶段,对虫体血淋巴内氨基酸的种类需求不同,其中赖氨酸、酪氨酸、精氨酸和丝氨酸等应该是白僵菌在桑天牛幼虫虫体内生长发育所需的主要氨基酸。
3 讨论昆虫血淋巴含有多种蛋白质,不仅与组织形成和物质代谢有关,而且与虫体生长发育的激素调控以及产生抗药性和免疫机制等有密切关系。血淋巴蛋白质的变化是昆虫新陈代谢正常与否的一个指标,通过对感病昆虫血淋巴蛋白的研究可以了解白僵菌在寄主组织内增殖时血淋巴中蛋白质的变化(陈长琨,1993)。
试验结果表明:处理2天时桑天牛幼虫血淋巴内蛋白质含量明显升高,可能是由于白僵菌的侵染,刺激了昆虫体内的防御系统,血淋巴中产生大量与免疫机制相关的蛋白质和酶类; 随后由于白僵菌在血淋巴中的大量增殖,昆虫各组织器官功能受阻,对食物消化和吸收功能减弱,血液中蛋白合成与转运受到抑制,同时由于组织被破坏,组织解离释放出更多蛋白而出现上述结果,这与该菌对桑天牛幼虫的入侵过程是一致的。5天后,体内蛋白质含量呈现持续下降,直至昆虫死亡。分析其原因,主要有以下几点:1)由于白僵菌感染,抵制了桑天牛幼虫对食物的消化和利用,其正常的代谢活动和生长发育受到影响,体内的蛋白质合成也受到阻碍; 2)白僵菌在昆虫血腔内增殖,消耗了虫体内大量蛋白质; 3)桑天牛需要消耗自身的营养物质(包括蛋白质)来维持基本的代谢活动; 且感染后期,由于其自身免疫调节功能的逐步丧失和白僵菌体外分泌酶等原因,使得宿主的可溶性和非可溶性蛋白质降解(陈伍国,2002)。
电泳试验中观察到:白僵菌感染后,血淋巴蛋白质图谱出现了明显的变化,蛋白质谱带在数量、颜色及宽度的变化与蛋白质含量的变化趋势相同,亦与白僵菌侵入过程一致。侵染第1天出现的b带可能是幼虫应激反应产生的免疫性蛋白或相关酶类,也可能是白僵菌产生的毒素蛋白或菌体自身蛋白的存在,其来源和生理功能需进一步研究。a带和c带在第5天开始减弱则是由于桑天牛幼虫受侵染后期正常的代谢活动遭到破坏,体内的蛋白质合成受阻,而桑天牛还需消耗自身的蛋白质来维持基本的代谢活动所致。第5天和第6天新增加的带可能是感染后期幼虫一些组织被破坏,组织解离释放出的新蛋白; 也可能是由于昆虫对食物的消化和吸收功能的减弱,致使血液中蛋白合成与转运受到抑制而滞留于血淋巴中的缘故。
白僵菌对各种昆虫毒力的高低,取决于昆虫体液成分能否满足该菌株对营养的需求,一般所谓敏感昆虫,就是其体液成分能符合菌株对营养的需求,否则毒力弱(吴振廷,1988)。桑天牛幼虫体液中含有丰富的营养成分,其中所含的游离氨基酸就达17种之多,能满足白僵菌的需求,在染病幼虫体液中常可见大量菌丝段,说明桑天牛幼虫较适合该菌的发育,白僵菌对其表现出较强的毒力。
感染24 h,游离氨基酸含量下降,主要是由于球孢白僵菌入侵寄主后,利用虫体内的养分进行营养生长。感染96 h,虫体内游离氨基酸含量逐渐上升,主要与自身免疫调节功能的逐步丧失以及组织被破坏、解离、蛋白质解体有关。说明白僵菌侵染桑天牛幼虫致死的过程中,不同发育阶段对寄主血淋巴中氨基酸的吸收和利用存在着数量差异,这与白僵菌在寄主血淋巴中增殖对营养物质的需求变化情况相一致,同时也与寄主的免疫反应及组织被破坏、蛋白质解体等有关。但总体上游离氨基酸含量的变化总是朝着有利于菌体生长发育的方向进行。侵染后期氨基酸含量的增加又促进了白僵菌在虫体内的大量增殖,以致氨基酸被全部吸收,虫体因出现“生理饥饿”而逐渐死亡。
纵观白僵菌对桑天牛幼虫的入侵致病过程可以看出,接种后24 h,孢子开始穿透体壁进入血腔,首先利用虫体的营养成分进行繁殖,因此游离氨基酸含量明显下降; 之后由于昆虫的免疫机制被激活,刺激产生相关的蛋白和酶类,使得蛋白质含量开始升高。但这种免疫作用并不能完全消除白僵菌菌丝在血腔内增殖,而随着菌丝在桑天牛幼虫血腔内的大量增殖和昆虫各器官功能逐渐丧失,昆虫的免疫机制被瓦解,组织被破坏,蛋白也被逐渐解体,以至于到侵染的后期血淋巴蛋白质含量开始下降,而氨基酸含量则明显升高。因此认为,血淋巴蛋白质和游离氨基酸含量在白僵菌入侵过程中发挥着重要的免疫作用,但这种免疫机制还有待进一步研究。
| [] | 陈长琨. 1993. 昆虫生理生化实验. 北京, 农业出版社. |
| [] | 陈伍国, 骆启桂, 王丹, 等. 2002. 中华卵索线虫感染棉铃虫后的血淋巴病理学研究. 华中师范大学学报:自然科学版, 36(3): 349–352. |
| [] | 丁德贵, 李增智, 樊美珍, 等. 2004. 球孢白僵菌种群在松林中的寄主转移及遗传多样性对松毛虫持续控制的影响. 应用生态学报, 15(12): 2315–2320. DOI:10.3321/j.issn:1001-9332.2004.12.023 |
| [] | 傅丽君. 2000. 小菜蛾感染球孢白僵菌后病理反应研究. 福建农业大学硕士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-ANHU804.001.htm |
| [] | 付丽君, 尤民生, 戴小华. 2001. 小菜蛾感染球孢白僵菌后血淋. 巴游离氨基酸的测定.华东昆虫学报, 10(1): 34–38. |
| [] | 郭志红, 崔永三, 杨弘平, 等. 2001. 卵孢白僵菌防治苗圃地蛴螬的应用技术. 东北林业大学学报, 29(6): 32–35. |
| [] | 赫英伟. 2010. 白僵菌的研究概况. 黑龙江农业科学(4): 145–147. |
| [] | 李庚祥, 陈斌, 李正跃, 等. 2009. 三株丝孢虫生真菌对松墨天牛幼虫的感染效应. 云南农业大学学报, 24(1): 47–50, 66. |
| [] | 李会平. 2007. 桑天牛幼虫高致病性白僵菌菌株筛选及其致病机理研究. 河北农业大学博士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-LYKE605.009.htm |
| [] | 李会平, 黄大庄, 王晓红, 等. 2006. 用黄粉虫诱集法分离球孢白僵菌及对桑天牛幼虫高毒力菌株的筛选. 蚕业科学, 43(11): 66–71. |
| [] | 林庆源. 2005. 荔枝蝽白僵菌优良菌株的筛选及应用技术的研究. 林业科学研究, 18(2): 143–146. |
| [] | 刘洪剑, 朴春根, 汪来发, 等. 2007. 白僵菌和肿腿蜂对松墨天牛幼虫的作用. 林业科学, 43(5): 64–68. |
| [] | 刘洪剑, 束庆龙, 汪来发, 等. 2009. 白僵菌孢悬液添加增效剂对松墨天牛幼虫的致死作用. 林业科学研究, 22(5): 652–656. |
| [] | 陆秀君, 李瑞军, 董立新, 等. 2008. 美国白蛾高毒白僵菌菌株筛选及其与高效氯氰菊酯的相容性. 植物保护学报, 35(6): 575–576. |
| [] | 牛宇. 2005. 病原物白僵菌对油松毛虫致病性研究. 山西大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10108-2005114061.htm |
| [] | 孙鲁娟, 吴孔明, 郭予元. 2001. 不同温、湿度下白僵菌对棉铃虫幼虫的致病力. 昆虫学报, 44(4): 501–506. |
| [] | 王达, 苏筱雨, 黄大庄, 等. 2009. 桑天牛幼虫血淋巴对球孢白僵菌的防御反应. 林业科学, 45(4): 83–87. DOI:10.11707/j.1001-7488.20090414 |
| [] | 王滨. 2003. 白僵菌持续控制马尾松毛虫的生物多样性和生态学基础. 浙江大学博士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-AHLY200503041.htm |
| [] | 王晓红, 纪惠芳, 黄大庄, 等. 2008. 桑天牛幼虫感染白僵菌后的组织病理学研究. 蚕业科学, 34(1): 18–23. |
| [] | 王云滨, 董建臻, 宋入梅, 等. 2007. 甜菜夜蛾优良球孢白僵菌菌株的筛选. 中国农学通报, 23(5): 346–348. |
| [] | 吴振廷. 1988. 菜青虫感染苏云金杆菌HD-1血淋巴的生理变化. 昆虫知识, 25(3): 149–151. |
| [] | 西北农业大学. 1986. 基础生物化学实验指导. 西安, 陕西科学技术出版社. |
| [] | 徐均焕, 冯明光, 童贤明. 2003. 球孢白僵菌孢子萌发促进剂的筛选及其对杀蚜效果的增强作用. 植物保护学报, 30(1): 45–50. |
| [] | 薛皎亮, 牛宇, 谢映平. 2006. 油松毛虫感染白僵菌后体内蛋白质、酯酶和多酚氧化酶的变化. 应用与环境生物学报, 12(6): 814–818. |
| [] | 许文耀. 2006. 普通植物病理学实验指导. 北京, 科学出版社. |
| [] | 张爱文, 刘维真, 农向群. 1992. 白僵菌可湿性粉剂的研制. 生物防治通报, 8(3): 118–120. |
| [] | Blake R, Bextun E, Harlan G. 2002. Field application of bait-formulated Beauveria bassiana alginate pellets for biological control of the red imported fire ant. Biol Control, 31(4): 746–752. |
| [] | Felipe T, Mario Z, Raquel A. 2004. Pathogenicity of Beauveria bassiana. Florida Entomol, 87(4): 533–536. DOI:10.1653/0015-4040(2004)087[0533:POBBDH]2.0.CO;2 |
| [] | Shmazu M, Sato H. 1995. Microbial control of the pine sawyer, Monochamus alternatus by Beauveria bassiana. International Symposium on Pine Wood Nematode.Beijing, China: 128–137. |
| [] | St Leger R J, Screen S. 2001. Prospects for strain improvement of fungal pathogens of insects and weeds//St Leger R J. Fungal as Biocontrol Agents.London:CABI Publish House: 219–237. |