 2012, Vol. 48
2012, Vol. 48文章信息
- 何春霞, 孟平, 张劲松, 高峻, 孙守家
- He Chunxia, Meng Ping, Zhang Jingsong, Gao Jun, Sun Shoujia
- 基于稳定碳同位素技术的华北石质山区2种果农复合模式水分利用研究
- A study on water use of two fruit tree-wheat intercropping systems in the rocky hilly region of north China with stable carbon isotope technique
- 林业科学, 2012, 48(5): 1-7.
- Scientia Silvae Sinicae, 2012, 48(5): 1-7.
-
文章历史
- 收稿日期:2011-05-24
- 修回日期:2011-08-08
-
作者相关文章



农林复合系统通过林木和作物配置,能使物种间生态互补,充分利用水肥光热等资源,提高系统的生态及经济效益。但随着林木根系和冠幅增大,农林复合系统各植物组分之间可能存在资源竞争,导致作物减产,尤其在温带半干旱地区地下部分水分竞争更为突出(毛瑢等, 2009)。采用合理的树种选择和时空搭配等措施能降低林木-作物间强烈的水分、养分等竞争。复合农林系统是太行山林业生态工程建设的重要内容之一,果农复合模式是其中一大主要类型。该区气候干旱、土层瘠薄、灌溉条件较差,水资源短缺制约了农林业的持续发展。开展华北石质山区不同农林复合模式植物的水分利用关系研究,对该区发展节水高效、持续稳定的农林复合系统具有重要意义(孙守家等, 2010)。
稳定碳同位素技术是近年来植物生理生态学研究中揭示植物与环境相互作用的有力工具之一。碳同位素效应能反映植物的碳和水分关系,尤其是在水分受限的情况下(Farquhar et al., 1984)。植物水分利用效率(water use efficiency,WUE)是指植物生理活动过程中消耗水形成有机物质的基本效率,是评价植物水分利用和抗旱特性的一个重要指标(Bohn et al., 2002),能为不同生境合适树种的选择提供理论依据。已有很多研究(李吉跃等, 2002;Anyia et al.,2007)证实,植物在轻度和中度干旱胁迫下WUE显著增大。由于植物组织的碳是在一段时间(如整个生长期)内累积起来的,其稳定碳同位素组成(stable carbon isotope ratio,δ13C)可以指示截至采样时植物生活过程的平均WUE,其可靠和稳定性已得到充分证实(Farquhar et al., 1982; 1989;Anyia et al., 2007;Chen et al., 2010)。该方法只需采集少量植物材料,不受时间和地点限制,方便保存和测定,简化了WUE的测定过程(Farquhar et al., 1984);比传统的收获法(生长季内植物收获干质量与田间耗水量的比值来表示WUE)更准确(Pieters et al., 2008)、快速、简便和破坏性小(Leffler et al., 2001);也比常规的光合仪法(以光合和蒸腾之比来表示WUE)优越,克服了其只能进行短期及瞬时WUE研究很难与植物的最终生产力和田间WUE联系起来的缺点(Farquhar et al., 1989;王建林等, 2010)。采用稳定碳同位素技术研究林农复合系统的水分利用还较少见。
核桃(Juglans regia)-小麦(Triticum aestivum)和石榴(Punica granatum)-小麦是华北石质山区较为普遍发展的农林复合模式。本文在冬小麦不同生育期内采用稳定碳同位素技术研究了上述2种复合系统和单作小麦的植物组分的δ13C值,结合生物量和气象数据资料计算不同模式中植物组分的WUE和耗水量(water use,WU),以期找出优化的间作模式,为该区农林复合系统的水分管理提供理论依据,并可丰富稳定碳同位素技术的应用和农林复合系统水分利用的研究方法。
1 试验地概况试验地设在河南省济源市境内的黄河小浪底森林生态系统定位研究站内(35°01′ N,112°28′ E),试验区地处太行山南段南麓,属暖温带大陆性季风气候。全年日照时数为2 367.7 h,年日照率为54%,多年≥0 ℃平均活动积温为5 282 ℃。平均降水量641.7 mm,主要分布在6—9月(占全年的68.3%)。试验区土壤以石灰岩风化母质淋溶性褐色土为主,土层厚50~80 cm,pH值7.65~8.50,石砾含量10%~18%,有机质含量8.28~16.50 g·kg-1,速效氮含量21.4~80.0 mg·kg-1,有效磷含量2.77~ 8.16 mg·kg-1,速效钾含量77.44~97.98 mg·kg-1。
2 材料与方法 2.1 试验材料研究对象为水平梯田核桃-小麦(walnut tree intercropping wheat, WIW)和石榴-小麦(megranate tree intercropping wheat, MIW)间作系统和单作小麦(monoculture wheat, MW)。核桃和石榴梯田均是东西向长140 m、南北宽20 m。果树均种植于2006年初,林带行向为东西向,株行距为3 m × 4 m。核桃和石榴的平均株高、冠幅、地径分别为4.85 m,3.38 m,9.06 cm和2.70 m, 2.02 m和5.60 cm。小麦品种为‘焦豫6号’,于2009年10月中旬播种、2010年6月初收获,行距20 cm,整个生育期无灌溉补水措施。间作小麦距离核桃和石榴南北侧树木行距均为60 cm。单作小麦位于2个复合模式附近的梯田地块中,与间作小麦的管理措施相同。
2.2 试验小区布置2个间作模式中分别选择1株果树及其周围间作小麦作为1个试验小区,分别在距离核桃和石榴树行北侧60 cm(N60)和190 cm(N190),南侧60 cm(S60)处的行间对小麦进行取样,各设3个试验重复。
2.3 气象因子观测小气候自动观测系统位于2个复合模式和单作小麦中间位置1 m高度处,连续观测空气温度(Ta)和相对湿度(RH)(HMP45C,Vaisala, Finland)、降雨量(P)(RM Young,Inc)等气象数据,数据采集器为CR23X(Campbell公司, 美国)。
2.4 稳定碳同位素的测定于小麦返青期(seedling establishment, SE)、拔节期(Jointing, J)、扬花-灌浆期(flowering-filling,F-F)、成熟期(Maturing,M)4个时期对果树和小麦取样,用于测定各植物组分的δ13C值和计算WUE。其中果树分东南西北4个方位取样,每个方位取5~8片叶,在小麦返青、果树还未发芽阶段,取1年生枝条(去皮),而在小麦拔节、果树萌叶阶段,取嫩叶;小麦按照2.2中布置的采样点取样,取旗叶,每个样品为15株小麦的混合样。样品在70 ℃烘箱中烘干48 h,粉碎过80目筛,在中国林业科学研究院稳定同位素比率质谱实验室分析。取处理好的样品3~5 mg在元素分析仪(Thermo Scientific Flash1112 HT,美国)中高温氧化还原为CO2,质谱仪(DELTA V Advantage Isotope Ratio Mass Spectrometer)检测CO2中的C得到样品中δ13C值,测定精度为0.1‰。同时用小气泵取单作和复合系统中0.5 m高度的空气样品装入气体采样袋中用于分析空气的CO2浓度和δ13C值,每样品3个重复。空气CO2浓度用气相色谱仪(Agilent 6820)、火焰离子化检测器(FID)测定,分析精度为5‰;空气δ13C值也用质谱仪采用高温燃烧法测定。植物和空气δ13C值的测定以PDB(Pee Dee Belemnite)为标准,根据下面公式计算(Farquhar et al., 1982):

|
(1) |
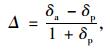
|
(2) |
式中:δ13C表示样品13C/12C与标准样品偏离的千分率,(13C/12C)PDB表示标准物质PDB的13C/12C。δa和δp分别为空气和植物材料的δ13C值。
2.5 小麦生物量、产量调查在小麦返青期、拔节期、扬花-灌浆期和成熟期4个时期测定小麦的生物量。小麦成熟后,在2.2中布置的每个取样点所在麦行各选取50 cm长小麦调查生物量、产量、收获指数HI(产量和生物量的比值)、千粒质量、单株麦粒数、株高、单位面积的麦株数和比叶质量(单位面积的叶片干质量leaf mass per area,LMA)等,将穗、根、茎、叶分别烘干测定生物量、δ13C值和碳含量(质谱仪法随同样品δ13C值同时测出)。
2.6 WUE等的计算根据Farquhar等(1982;1984;1989)的研究:
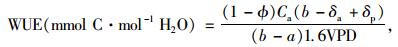
|
(3) |
式中:a,b分别为CO2扩散和羧化过程中的同位素分馏系数,其中a=4.4‰,b=27‰;Ca大气CO2浓度;数值1.6为水蒸汽和CO2在空气中的扩散比率。Φ为植物整个生长期叶片夜间呼吸和其他器官呼吸消耗掉的碳的比率,取Φ=0.3(Evans, 1983b)。VPD为叶片内外蒸汽压差(vapor pressure deficit),根据植物生长过程中取样日期前一段的平均白日(7:00—17:30)气象数据(气温、空气湿度等)计算得出(Abbate et al., 2004; Hu et al., 2010):
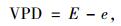
|
(4) |

|
(5) |
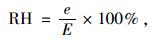
|
(6) |

|
(7) |
式中,T为叶片温度,根据Allen(2003),气温每升高1 ℃,叶片温度升高0.72 ℃;RH为大气相对湿度(relative humidity);0.611为t=0 ℃时纯水平面上的饱和水汽压;e为实际水汽压;E为同温度下的饱和水汽压。
同时,WUE又是植物在一段时间内同化的碳总量与总耗水量(water use,WU,g·m-2)的比值,可根据各个器官的生物量干质量(dry weight,DW,g)和含碳率(carbon content,CC,mg·g-1 DW)计算出干物质碳含量,即:
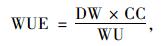
|
(8) |
将(3)式中WUE的单位mmol C·mol-1 H2O换算成mg C·g-1 H2O,代入公式(8)即计算出单位面积的实际耗水量WU。
2.7 数据分析采用EXCEL和SPSS18.0统计软件对数据进行分析。
3 结果与分析 3.1 气象因子如图 1所示, 2009—2010年冬小麦生育期内冬季日均温为2.28 ℃,生长季的平均气温为16.62 ℃。整个生育期降雨199.9 mm, 不到年平均降雨量的1/3。其中,播种至冬前期为77.4 mm,返青前45.7 mm,拔节期3.6 mm,孕穗至灌浆期57.6 mm,成熟期6.5 mm。RH的季节变化不明显,冬季稍低。日平均VPD的变化趋势与温度大概一致,冬季小、生长季大、雨天降低。

|
图 1 冬小麦生育期内气象因子变化 Fig.1 Changes of meteorological factors during winter wheat growth period |
核桃和石榴的δ13C和WUE分别平均为-25.777‰, -26.404‰和14.494, 13.642 mmol C·mol-1 H2O。在4月12号(小麦的拔节期)之前为核桃>石榴,且两者差异显著(均P=0.000);之后为石榴>核桃,但差异不显著(P=0.150和0.132)(图 2)。核桃和石榴的Δ值平均为15.229‰和15.863‰。2树种的δ13C都是在小麦拔节期的最大(均为P=0.000),核桃的季节变异最大(2.449‰),石榴较小(0.321‰)。而核桃和石榴的WUE的季节间变异分别为10.771和8.315mmol C·mol-1 H2O(P=0.000),均为3月最大,4月次之,5月最低。

|
图 2 核桃和石榴的δ13C,Δ和WUE差异 Fig.2 Difference of δ13C, Δ and WUE in J. regia and P. granatum tree SE:返青期Seedling establishment; J:拔节期Jointing; F-F:扬花-灌浆期Flowering-filling; M:成熟期Maturing.下同The same below. |
核桃间作小麦(WIW)、石榴间作小麦(MIW)和单作小麦(MW)的δ13C和WUE分别为-27.151‰,-26.788‰,-27.338‰和12.780,13.285,12.622 mmol C·mol-1 H2O。如图 3所示,模式间变异在小麦返青期分别为0.970‰和1.540 mmol C·mol-1 H2O,其中MIW>MW>WIW(P=0.000);在拔节期分别为1.317‰和1.471 mmol C·mol-1 H2O(P=0.000),WIW和MIW的WUE在拔节期分别比MW显著提高13.84%和10.86%;而在开花期和成熟期差异不显著(P=0.195,P=0.133)。MIW在4个生育期均比MW的WUE显著提高4.29%,10.86%, 3.01%和3.49%。

|
图 3 不同间作模式小麦的δ13C,Δ和水分利用差异 Fig.3 Difference of wheat δ13C, Δ and water use in different intercropping systems MW:单作小麦Monoculture wheat; WIW:核桃间作小麦Walnut intercropping wheat; MIW:石榴间作小麦Megranate intercropping wheat.下同The same below. |
单作小麦的δ13C为返青期>成熟期>扬花-灌浆期>拔节期(P=0.000)(图 3),WUE为返青期>扬花-灌浆期>成熟期>拔节期(P=0.000),2指标季节变异分别为1.351‰和9.203 mmol C·mol-1 H2O。而2个间作模式小麦WUE的季节差异显著(P=0.000):返青期>拔节期>成熟期>扬花-灌浆期(图 3)。
越往生长后期小麦需水量越多(除了单作小麦拔节期耗水多于扬花-灌浆期外),在返青期、拔节期、扬花-灌浆期和成熟期的耗水量分别占总耗水量的10.0%~10.4%,22.1%~27.2%,22.9%~27.7%和39.6%~39.9%,且生育期间差异均显著(均为P=0.000)(图 3)。其中在前3个生育期单作小麦的耗水量所占比例小于间作小麦,而在成熟期所占比例则大于间作小麦。WIW和MIW的总耗水量为185.92,175.27 mm,分别比单作小麦(253.81 mm)少耗水26.75%和30.94%,且在4个生育期均显著少于单作小麦(分别为P=0.014,0.006,0.046和0.050),表明间作系统显著降低了小麦的耗水量。
如图 4所示,位于核桃和石榴果树南北不同距离处间作小麦的WUE表现为:树木北侧N60,N190和南侧S60在4个生育期均差异不显著(P>0.05)。而耗水量是核桃N60的最多,S60的最少;石榴是S60最多,N190(树行中间)的最少。

|
图 4 果树不同距离处间作小麦的水分利用差异 Fig.4 Water use of wheat at different distance from intercropping fruit trees |
单作小麦的产量、总生物量和千粒质量都显著高于间作小麦的(P=0.001,P=0.033,P=0.006),分别比核桃间作小麦的高出26.79%,27.12%和14.87%,比石榴间作小麦高32.19%,28.53%和10.26%(表 1);核桃间作小麦的产量和生物量都高于石榴间作小麦,但差异不显著。而单作小麦比间作小麦的收获指数HI略低。
|
|

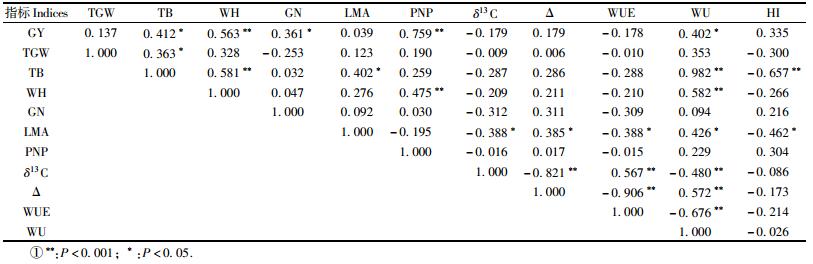
表 2的相关分析得出产量与麦株高、单位面积麦株数(均为P < 0.001)、总生物量、单株麦粒数、耗水量(P < 0.05)呈显著正相关,与千粒质量、LMA和Δ呈正相关、与δ13C和WUE呈负相关,但相关性均不显著。Anyia等(2007)对大麦(Hordeum vulgare)的研究也得出叶片的Δ与产量相关性不明显。总生物量与麦株高、耗水量(均为P < 0.001)、产量、千粒质量和LMA(P < 0.05)呈显著正相关,与HI显著负相关(P < 0.001);与单株麦粒数、单位面积麦株数和Δ呈正相关,与δ13C和WUE呈负相关,但相关性均不显著。
|
|
水分亏缺条件下植物会提高水分利用效率(Farquhar et al., 1984;Anyia et al., 2007)。间作小麦在返青期和拔节期的WUE显著高于单作小麦可能是由于在此时期小麦快速生长,而果树也正处于树液开始流动、发芽萌叶期,均需水较多,而这段时间雨水也较少,造成水分缺乏,相对单作小麦没有果树与其竞争水分,间作小麦与果树产生水分竞争。灌浆和成熟期间作小麦的WUE和单作小麦差异不明显,除了这一时期降雨稍多,补充了水分外,另一个重要原因可能是此时果树已经形成树冠遮荫,可有效地减少地表水分散失、增加空气湿度、降低叶温、气温以及VPD,改善间作系统的小气候和土壤水分状况,这些都将减轻水分亏缺对林冠下作物的影响(Quero et al., 2006),在生长后期单作小麦所消耗水分比例大于间作小麦的总耗水比例也充分证实了这一点。
在整个小麦生长季,WUE都以石榴间作系统的最大,核桃间作小麦在返青期和拔节期的WUE也大于单作小麦,并且位于核桃和石榴不同距离处间作小麦的WUE和总耗水量无明显差异,这可能是4年生株行距3 m × 4 m的核桃和石榴根系已经布满整个间作系统,与间作小麦在各个位置都有地下部分生态位重叠,并存在水分竞争。
虽然单作小麦的单株麦粒数和单位面积的小麦株数与2个间作模式差异不显著,但千粒质量和产量却显著高于间作模式小麦的,可能是间作果树的水分竞争或者遮荫影响了小麦灌浆和籽粒饱满,减小了麦粒质量和小麦产量,这也需要进一步的研究。产量和生物量都与耗水量呈正相关,但与WUE相关性不显著表明小麦形成的生物量和产量越多需要吸收和消耗的水分也越多,证实了水分亏缺条件下的小麦高WUE是以绝对产量(生物量)减少为代价的(Bloch et al., 2006;Chen et al., 2010)。间作小麦的HI比单作小麦略高可能是间作提高了小麦对麦粒(产量)的投入。HI与LMA呈负相关也暗示了投入到叶片中的碳越少则用于形成产量的碳越多。
总的来说,株行距3 m × 4 m的核桃-小麦和石榴-小麦间作模式在果树的第4年,核桃和石榴的单株果树产量平均分别为0.88,14.67 kg,与清耕果园的产量(0.94,14.73 kg)均差异不显著(P=0.829,P=0.988);间作小麦比单作小麦分别减产26.79%和32.19%,同时减少了26.75%和30.94%的水分消耗,间作系统显著降低了小麦的耗水量,提高了水分利用效率,尽管减产但仍有较好的作物收益,并与清耕果园相比对果树影响很小。因此应继续发展果农间作系统,增加作物收获和经济效益。在果树完全郁闭后可以考虑种植耐荫作物或药材等,以最大限度地提高复合系统的经济效益。
| [] | 李吉跃, 周平, 招礼军. 2002. 干旱胁迫对苗木蒸腾耗水的影响. 生态学报, 22(9): 1380–1386. |
| [] | 毛瑢, 曾德慧. 2009. 农林复合系统植物竞争研究进展. 中国生态农业学报, 17(2): 379–386. |
| [] | 孙守家, 孟平, 张劲松, 等. 2010. 华北石质山区核桃-绿豆复合系统氘同位素变化及其水分利用. 生态学报, 30(14): 3717–3726. |
| [] | 王建林, 杨新民, 房全孝. 2010. 不同尺度农田水分利用效率测定方法评述. 中国农学通报, 26(6): 77–80. |
| [] | Abbate P E, Dardanelli J L, Cantarero M G, et al. 2004. Climatic and water availability effects on water-use efficiency in wheat. Crop Science, 44(2): 474–484. DOI:10.2135/cropsci2004.4740 |
| [] | Allen L H, Pan Jr D, Boote K J, et al. 2003. Carbon dioxide and temperature effects on evapotranspiration and water use efficiency of soybean. Agronomy Journal, 95(4): 1071–1081. DOI:10.2134/agronj2003.1071 |
| [] | Anyia A O, Slaski J J, Nyachiro J M, et al. 2007. Relationship of carbon isotope discrimination to water use efficiency and productivity of barley under field and greenhouse conditions. Journal of Agronomy & Crop Science, 193(5): 313–323. |
| [] | Bloch D, Hoffmann C M, Marlander B. 2006. Impact of water supply on photosynthesis, water use and carbon isotope discrimination of sugar beet genotypes. European Journal of Agronomy, 24(3): 218–225. DOI:10.1016/j.eja.2005.08.004 |
| [] | Bohn B A, Kershner J L. 2002. Establishing aquatic restoration priorities using a watershed approach. Journal of Environmental Management, 64(4): 355–363. |
| [] | Chen J, Chang S X, Anyia A O. 2011. The physiology and stability of leaf carbon isotope discrimination as a measure of water-use efficiency in barley on the canadian prairies. Journal of Agronomy & Crop Science, 197(1): 1–11. |
| [] | Evans J R. 1983. Nitrogen and photosynthesis in the flag leaf of wheat(Triticum aestivum L.). Plant Physiology, 72(3): 297–302. |
| [] | Farquhar G D, Ehleringer J R, Hubick K T. 1989. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 40(1): 503–537. DOI:10.1146/annurev.pp.40.060189.002443 |
| [] | Farquhar G D, O'Leary M H, Berry J A. 1982. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, 9(2): 121–137. DOI:10.1071/PP9820121 |
| [] | Farquhar G D, Richards R A. 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Australian Journal of Plant Physiology, 11(6): 539–552. DOI:10.1071/PP9840539 |
| [] | Hu J, Moore D J P, Riveros-Iregui D A, et al. 2010. Modeling whole-tree carbon assimilation rate using observed transpiration rates and needle sugar carbon isotope ratios. New Phytologist, 185(4): 1000–1015. DOI:10.1111/j.1469-8137.2009.03154.x |
| [] | Leffler A J, Evans A S. 2001. Physiological variation among Populus fremontii populations: short-and long-term relationships between δ13C and water availability. Tree Physiology, 21(15): 1149–1155. DOI:10.1093/treephys/21.15.1149 |
| [] | Pieters A J, Nunez M. 2008. Photosynthesis, water use efficiency and δ13C in two rice genotypes with contrasting response to water deficit. Photosynthetica, 46(4): 574–580. DOI:10.1007/s11099-008-0096-7 |
| [] | Quero J L, Villar R, Marañón T, et al. 2006. Interactions of drought and shade effects on seedlings of four Quercus species: physiological and structural leaf responses. New Phytologist, 170(4): 819–834. DOI:10.1111/nph.2006.170.issue-4 |