 2012, Vol. 48
2012, Vol. 48文章信息
- 周建云, 李荣, 张文辉, 何景峰
- Zhou Jianyun, Li Rong, Zhang Wenhui, He Jingfeng
- 不同间伐强度下辽东栎种群结构特征与空间分布格局
- Effects of Thinning Intensity on Structure Characteristics and Spatial Distribution of Quercus wutaishanica Populations
- 林业科学, 2012, 48(4): 149-155.
- Scientia Silvae Sinicae, 2012, 48(4): 149-155.
-
文章历史
- 收稿日期:2011-05-17
- 修回日期:2011-10-14
-
作者相关文章



2. 石河子大学农学院 石河子 832000
2. Agriculture College of Shihezi University Shihezi 832000
种群结构是植物种群的重要属性,反映种群个体在空间上的组配方式和发展趋势;种群分布格局是指种群个体在水平空间的配置状况或分布状态,反映种群个体在水平空间上彼此间的相互关系(蒋雪琴等,2009)。研究群落内优势种的种群结构和空间分布格局,有助于分析种群演替动态规律(Masaki et al., 2004),了解种群的生物学特性及其与环境因子之间的相互关联(Dray et al., 2006;张健等, 2007;惠刚盈等, 2007),对种群恢复具有要意义。
辽东栎(Quercus wutaishanica)林是黄土高原地区典型的地带性植被,辽东栎的持续发育,将对当地的生态防护和天然林持续发育起着举足轻重的作用(李荣等,2011a)。近几十年来,发挥森林自我更新能力的近自然林经营方式成为森林经营的主要方式之一,它既能最大限度地保护生物多样性,提升森林生态功能,又能产生一定的经济效益,因而受到国内外同行的认可(Parrotta et al., 1997;Olsthoorn et al., 1999;李荣等,2011a)。此前有关辽东栎种群的研究主要集中在种群动态(王巍等,1999;卢彦昌等,2006;冯云等,2007)、物种多样性(赵则海等,2002;康永祥等,2007)和空间分布格局(王巍,2000;冯云等,2009)等方面,对种群结构特征和空间分布格局方面的研究主要侧重某一地区或者同一地区不同森林类型上(张文辉等,2002;伊力塔等,2008;胡秀娟等,2010),有关间伐对辽东栎种群结构特征和空间分布格局的影响研究未见报道。
本研究以黄土高原地区黄龙山林区近自然经营间伐6年后的辽东栎林为对象,从龄级结构、静态生命表、存活曲线和种群空间分布格局等方面,研究不同间伐强度(封禁为对照,间伐材积强度分别为13.4%和30.0%)对辽东栎种群结构特征与空间分布格局的影响,为合理经营和恢复辽东栎林提供依据。
1 研究区概况研究区域选择在陕北黄土高原延安市黄龙山林业局蔡家川林场,地理位置为109°38′49″—110°12′47″ E,35°28′46″—36°02′01″ N,海拔962.6~1 783.5 m,属暖温带半湿润与半干旱气候的过渡地带,年平均气温8.6 ℃,最高气温36.7 ℃,最低气温-22.5 ℃,无霜期126~186天,年均降水量611.8 mm。地带性植被为暖温带落叶阔叶林,以辽东栎、油松(Pinus tabulaeformis)、茶条槭(Acer ginnala)、山杨(Populus davidiana)和白桦(Betula platyphylla)为建群种形成的纯林或者混交林呈镶嵌性分布(李荣等,2011a)。
该地区大部分天然辽东栎林林龄在50年左右,乔木层郁闭度0.80~0.90,林下有树木自然枯死现象。2004年底,选择辽东栎占林冠层80%以上的辽东栎天然次生林,以近自然经营理念为指导,设计间伐材积强度为13.4%(弱度间伐)和30.0%(强度间伐)的林分各10.0 hm2,实施目标树单株林分作业,原则是间密留匀,留优去劣,使林木分布均匀,林分结构更为合理。2004年底实施采伐作业,间伐后林地郁闭度在0.6以上(李荣等,2011a)。
2 研究方法 2.1 样地设置与调查2010年8月(伐后第6年),设20 m×20 m的样地12个,其中未间伐区(对照)、13.4%间伐区(弱度)和30.0%间伐区(强度)各4个(表 1)。在每块样地(20 m×20 m)中采用相邻格子法设置5 m×5 m的小样方。以样地的一条边为x轴,以其偏东向的垂直边作为y轴,记录每一株辽东栎的坐标值,同时每木检尺,记录其高度和胸径(或者基径)。
|
|
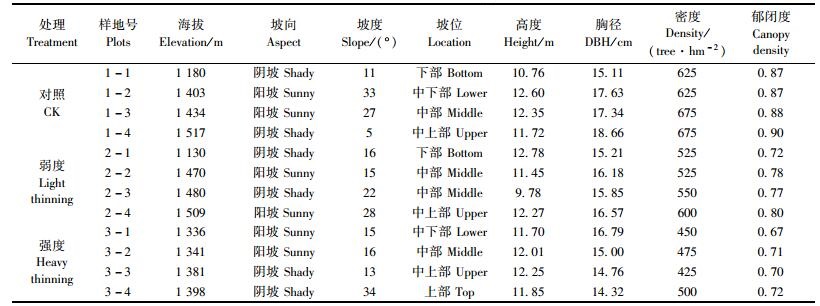
辽东栎种群起源有实生和萌生2种途径。在幼苗幼树阶段,萌生苗在空间分布、生长速度和对空间资源的利用等方面均占有明显的优势,龄级以实际年龄来划分不够合理(高贤明等,2001)。本研究以个体高度(幼苗幼树)及胸径(径级)代替年龄结构来分析种群动态。将辽东栎种群划分为4个生长阶段:幼苗(Ⅰ级)、幼树(Ⅱ~Ⅲ级)和小树(Ⅳ~Ⅶ级)和大树(Ⅷ~Ⅹ级)(高贤明等,2001;张赟等,2008)。幼苗幼树龄级划分标准为:Ⅰ级(高度H≤50 cm)、Ⅱ级(50 cm<H≤100 cm)、Ⅲ级(H>100 cm,DBH<4 cm)3个级别;根据树木的胸径,从4 cm开始,胸径每增加4 cm为一个径级,即Ⅳ级(4 cm≤DBH<8 cm)、Ⅴ级(8 cm≤DBH<12 cm)、Ⅵ级(12 cm≤DBH<16 cm)、Ⅶ级(16 cm≤DBH<20 cm)、Ⅷ级(20 cm≤DBH<24 cm)、Ⅸ级(24 cm≤DBH<28 cm)和Ⅹ级(DBH≥28 cm)(高贤明等,2001)。将龄级从小到大的顺序看作是时间顺序关系,统计各龄级株数,编制种群静态生命表(陈远征等,2006;郝朝运等,2009),并绘制存活曲线。
2.3 种群空间分布格局类型以样地内辽东栎个体定位数据为依据,应用相邻格子法,分5 m×5 m,5 m×10 m,10 m×10 m的样方格子分别进行统计分析;应用离散分布理论拟合,χ2检验判定与泊松分布、负二项分布、奈曼A型分布的吻合程度。聚集强度的测定采用负二项参数K、扩散系数(C)、扩散指数(Iδ)、Cassie指数(1/K)、丛生指标(I)和聚块性指标(m*/m)(王成磊等,2011)。
3 结果与分析 3.1 不同间伐强度下辽东栎种群的龄级结构不同间伐强度下辽东栎种群年龄结构呈现幼苗幼树比例最大,大树比例次之,小树比例最小的规律(图 1)。辽东栎幼苗幼树(Ⅰ~Ⅲ龄级)数量均较多(在对照、弱度和重度间伐样地,分别占全部个体数量的67.70%,74.64%和77.17%),表明辽东栎幼苗幼树数量足够使种群进行自我更新。与对照样地相比,弱度间伐和强度间伐样地中Ⅷ~Ⅹ龄级大树因间伐而数量分别降低29.03%和35.48%,而Ⅰ~Ⅲ龄级幼苗幼树数量分别增加18.81%和30.28%,Ⅳ~Ⅴ龄级小树数量分别增加4.76%和9.52%,说明间伐措施改善林内光照等生境条件,促进林下幼苗幼树和小树数量增加。

|
图 1 不同间伐强度下辽东栎种群林龄级结构 Fig.1 Age class of Q. wutaishanica population in different treatments |
由于不同间伐强度下林地生境条件的差异,辽东栎种群在期望寿命、死亡率、消失率等动态参数方面略有差异,但各参数在不同间伐强度下的总趋势较为相似。种群期望寿命反映个体的平均生存能力,呈现“低—高—低”的“纺锤型”模式。种群死亡率在Ⅰ~Ⅲ龄级最高,随着龄级的增加,死亡率呈现“高—低—高”的“哑铃型”模式(表 2)。
|
|
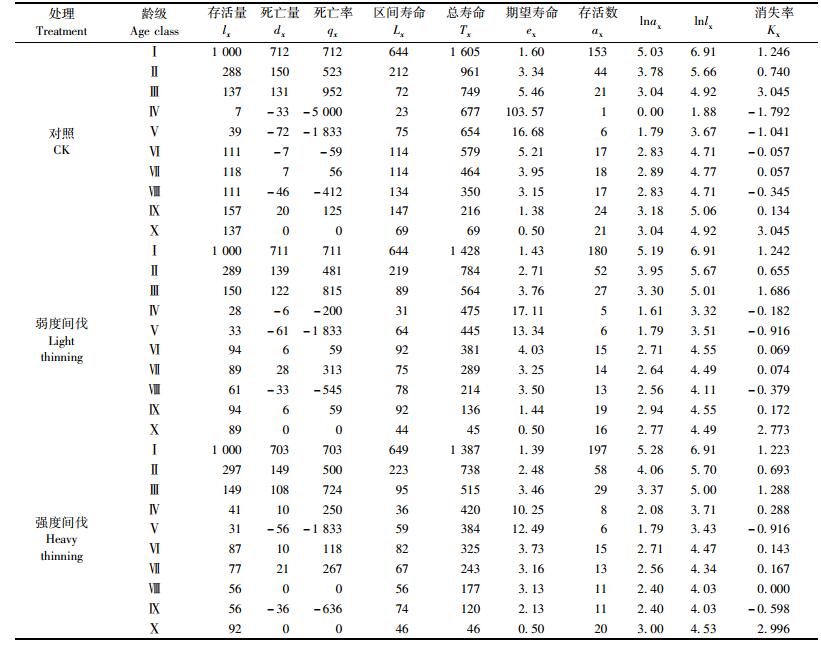
辽东栎种群的存活曲线接近于DeeveyⅢ型,即幼苗幼树期(Ⅰ~Ⅲ龄级)死亡率很高,种群个体数量急剧减少(图 2)。说明辽东栎幼年受环境影响和种间、种内竞争,导致较高的死亡率;随着年龄增加,死亡率逐渐下降,种群规模趋于稳定,种群实现了定居,死亡率维持在较低水平, 直至生理衰退年龄。

|
图 2 不同间伐强度下辽东栎种群的存活曲线 Fig.2 Survival curve of Q. wutaishanica population in different treatments |
不同间伐强度下辽东栎种群的存活曲线略有差异(图 2),间伐措施促进种群幼苗更新。3种处理在Ⅰ~Ⅲ龄级,幼苗存活率差异较小,对照样地略低于间伐样地;Ⅳ~Ⅴ龄级小树存活率较低,且对照<弱度<强度,说明此阶段为辽东栎种群发育的瓶颈,尤其是对照样地,此阶段小树数量明显不足;Ⅵ~Ⅹ龄级阶段存活曲线较为平滑稳定,且对照>弱度>强度,说明辽东栎达到Ⅵ龄级以上,存活率较高。在对照样地和弱度间伐样地,Ⅸ~Ⅹ龄级存活率较高于Ⅵ~Ⅷ龄级。
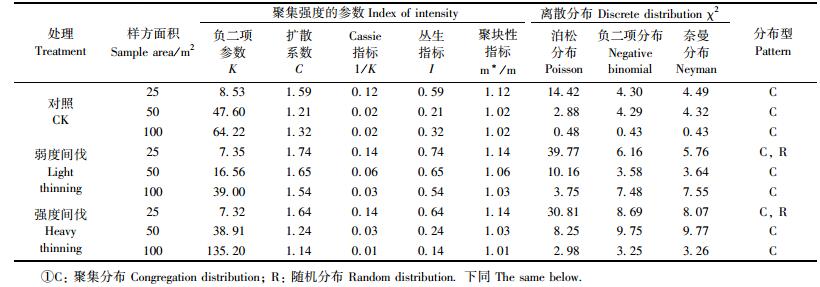
3.4 不同间伐强度和取样面积下辽东栎种群分布格局按3种大小格子来分析辽东栎种群的空间分布类型和聚集强度(表 3),辽东栎种群均呈聚集分布,种群格局强度随取样尺度由25~100 m2,聚集强度逐渐降低。不同间伐强度下辽东栎种群各项聚集指标略有差异,对照样地在所有尺度下均为聚集分布;弱度间伐和强度间伐样地在25 m2呈现出聚集分布和随机分布。
|
|
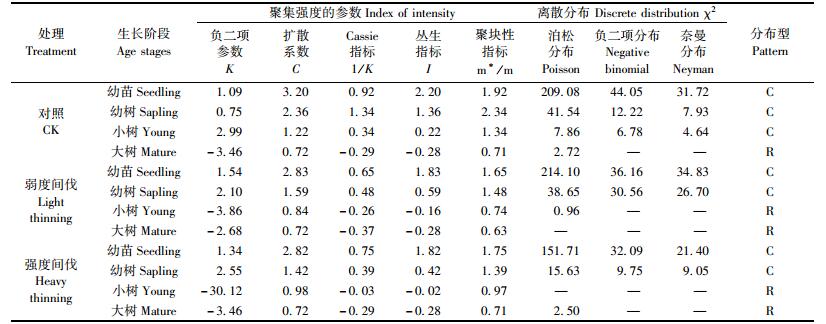
将辽东栎划分为幼苗、幼树、小树和大树4个阶段,在25 m2尺度下分析其种群分布格局,在不同强度间伐下,辽东栎种群各项聚集指标略有差异(表 4)。幼苗、幼树阶段个体多呈聚集分布,小树和大树阶段个体一般呈随机分布;在从幼苗—大树的时间序列上,种群的分布格局变化趋势是从聚集到随机。
|
|
在黄龙山林区,辽东栎的龄级结构是幼苗幼树比例最大、大树比例次之、小树比例最小。无论在间伐林地,还是在封育林地,林下辽东栎幼苗幼树(Ⅰ~Ⅲ龄级)数量均较多,说明该种群能够进行自我更新,种群具有较强的持续发育潜力。辽东栎幼苗幼树储备丰富,主要原因在于辽东栎能产生大量的、高质量的、广泛扩散的种子(魏瑞等,2009),为幼苗的产生提供基础。幼苗的大量产生必然会竞争有限的环境资源,在竞争中部分幼苗死亡,从而使得幼苗转变成幼树的死亡率很高。随着年龄增长,种群的抗性及适应环境的能力随之增强,死亡率也逐渐趋于稳定(王巍等,2000)。间伐后种群密度减小,林内光照增强,林内保留母树结实量增加,同时大大提高土壤的温度,种子落入土壤并萌发的机会增多,促进种子萌芽与幼苗生长,使种群幼苗幼树数量增加(李荣等,2011a)。
随着龄级的增加,辽东栎死亡率在大体上呈现“高—低—高”的“哑铃型”模式,种群期望寿命呈现“低—高—低”的“纺锤型”模式。说明辽东栎种群在其生长过程中,种群幼苗幼树的数量较大,表现出丰富的后备资源,但幼苗幼树易受恶劣环境与动物取食的影响,很难顺利转变成小树(张知彬,2001;Li et al., 2003)。虽然林内小树的数量不多,但辽东栎具有很强的萌生能力,萌生苗作为种群繁衍和稳定的一种途径,可迅速扩大幼树群体,使其顺利克服种群更新瓶颈,解决了辽东栎种群更新过程中“更新苗匮乏”的现象(高贤明等,2001)。小树一旦长成大树,其存活率就有所增加。辽东栎幼苗幼树在高度郁闭的林中处于“坐待”状态(李荣等,2011b),一旦间伐,林窗打开,幼苗幼树会快速生长,占领林地空间(王巍等,2000)。因此,间伐措施促进辽东栎小树(Ⅳ~Ⅶ龄级)数量增加。
辽东栎种群的存活曲线接近于DeeveyⅢ型。Ⅸ-Ⅹ龄级存活率较高于Ⅵ~Ⅷ龄级,原因在于辽东栎属于喜阳植物,在郁闭度较高的样地中,占据最高层空间的辽东栎(Ⅸ~Ⅹ龄级)可能会抑制较低层(Ⅵ~Ⅷ龄级)辽东栎的生长。研究区辽东栎林为近成熟林,郁闭度较高,林冠下没有充足的光照,辽东栎林地处在缓慢退化的状态。间伐后,林地光照、养分等增大,处在林冠下层的辽东栎存活率增高,促使种群结构优化、林木质量提高(李荣等,2011b)。
4.2 辽东栎种群分布格局辽东栎种群的空间分布格局受群落内小环境及辽东栎树种本身的生物学特性的共同影响(伊力塔等,2008)。本研究中,辽东栎分布格局从整体上看为聚集分布,从而形成了种群斑块镶嵌分布。这与伊力塔等(2008)、胡秀娟等(2010)和李宗峰等(2007)研究辽东栎种群空间分布格局相类似,他们都认为聚集分布可以增强辽东栎种群的竞争能力。这种种群聚集分布的研究结果与其他天然种群相类似(Toriola et al., 1998;Rigg,2005;缪宁等,2008)。
不同发育阶段辽东栎的聚集强度有差异。这种不同年龄阶段存在的空间格局差异,与种群新生个体的产生、成年个体的死亡以及外界的干扰有关(蒋雪琴等,2009)。在25 m2尺度上,幼苗幼树呈强烈的聚集分布,原因在于辽东栎存在有性生殖与无性生殖(李荣等,2011b)。一方面,母树不仅提供了种源,而且还创造了有利于种子萌发生长的小环境,故实生幼苗幼树多散布在母树周围,使得种群的繁衍顺利进行(Haase,2001);另一方面,辽东栎萌生力强,以伐根萌蘖繁殖为主,常在母树周围形成幼树期的聚集分布(胡秀娟等,2010)。幼苗幼树的聚集分布,有利于其相互庇护,共同抵御外界环境,提高种群存活机会(缪宁等,2008)。间伐样地中辽东栎幼苗幼树聚集度高于对照样地,原因在于本研究采用近自然经营间伐,实施目标树单株林分作业,同时保留了大量优质辽东栎母树(李荣等,2011c),林冠层出现空间上镶嵌的斑块状空隙更加有利于间伐样地中的幼苗幼树呈强烈聚集分布。
在一定尺度范围中,辽东栎个体在小树和大树阶段一般呈随机分布,这与伊力塔等(2008)认为随着年龄增长,辽东栎种群的分布格局变化趋势是从聚集到随机分布的研究结果相一致。这在其他栎类研究中也有相似的结论,如巨天珍等(2005)研究锐齿栎(Quercus aliena var. acuteserrata),张文辉等(2002)研究栓皮栎(Quercus variabilis),他们均认为随着年龄的增加,种内与种间竞争加剧,种群密度下降,种群一般由聚集向随即分布转变。其他天然群落的格局动态研究结果也表明:随着龄级的增大,种群的聚集程度逐渐降低,这有利于个体获得足够的资源(Hou et al., 2004;缪宁等,2008;王成磊等,2011)。
4.3 不同间伐措施对辽东栎种群结构与分布格局的影响研究区域辽东栎林是黄土高原地区比较稳定的森林植物群落(李荣等,2011a),即使在未采伐(对照)区, 林下也有少量辽东栎幼苗幼树,说明在当地辽东栎天然更新没有困难。但是在本研究中,未间伐区辽东栎林郁闭度较高,林冠下幼树数量明显不足,导致辽东栎林地处在缓慢退化的状态。仅靠自然恢复促使森林结构优化、森林林分质量提高和生态效益提高等都过于缓慢。近自然经营是一种人力与自然力结合在一起,人力推动自然力,最大限度发挥自然力的作用,促使森林可持续发展的贴近自然的森林经营模式(李荣等,2011c)。近自然经营(弱度或强度)间伐使种群成树数量减少后,不仅促进其幼苗幼树数量增加,还促进林冠中层的小树数量增加,导致种群结构更加合理,分布格局更加优化,加快了更新速度。综合当前不同间伐措施对辽东栎种群结构与分布格局的影响,结果表明强度间伐更有利于辽东栎种群的更新。
5 结论在不同取样尺度条件下,辽东栎种群随年龄增大聚集强度呈现减小趋势;种群在幼龄阶段呈较强的聚集分布,随年龄增长种群趋向随机分布;间伐样地种群聚集强度高于对照样地,表明了辽东栎喜阳的生物学特性,随着年龄增长需光性增强,特别是进入生殖期,光照条件影响结实质量和数量;随着间伐强度增加,种群结构更为合理,群落状态趋于稳定。辽东栎在25 m2尺度上高度聚集生长,这个尺度也可能是种群自然更新的最适面积。在森林抚育中,森林经营作业面积应尽量接近聚集性最强的面积尺度,充分利用自然因素的耦合效应,更加有利于种群恢复。
从整体来看,辽东栎种群能以幼苗幼树丰富的储备量、较强的萌生能力以及成年树木较强的存活率在群落中保持稳定的优势,表现出其对环境较强的适应能力,这也是辽东栎长期稳定存在并成为优势群落的原因。采伐强度30.0%(强度间伐)更能促进辽东栎群落结构优化,利于辽东栎的生长与更新。
| [] | 陈选征, 马祥庆, 冯丽贞, 等. 2006. 濒危植物沉水樟的种群生命表和谱分析. 生态学报, 26(12): 4267–4272. DOI:10.3321/j.issn:1000-0933.2006.12.046 |
| [] | 冯云, 马克明, 张育新, 等. 2007. 北京东灵山辽东栎(Quercus liaotungensis)林沿海拔梯度的物种多度分布. 生态学报, 27(11): 4743–4750. DOI:10.3321/j.issn:1000-0933.2007.11.043 |
| [] | 冯云, 马克明, 张育新, 等. 2009. 北京东灵山地区辽东栎(Quercus liaotungensis)种群生活史特征与空间分布. 生态学杂志, 28(8): 1443–1448. |
| [] | 高贤明, 王巍, 杜晓军, 等. 2001. 北京山区辽东栎林的径级结构、种群起源及生态学意义. 植物生态学报, 25(6): 673–678. |
| [] | 郝朝运, 张小平, 张昱, 等. 2009. 濒危植物永瓣藤的种群生命表与动态分析. 林业科学, 45(9): 79–84. DOI:10.11707/j.1001-7488.20090914 |
| [] | 胡秀娟, 程积民, 杨晓梅, 等. 2010. 黄土高原子午岭林区辽东栎种群分布格局及群落特征研究. 西北林学院学报, 25(5): 1–6. |
| [] | 惠刚盈, 李丽, 赵中华, 等. 2007. 林木空间分布格局分析方法. 生态学报, 27(11): 4717–4728. DOI:10.3321/j.issn:1000-0933.2007.11.040 |
| [] | 蒋雪琴, 刘艳红, 赵本元. 2009. 湖北神农架地区巴山冷杉(Abies fargesii)种群结构特征与空间分布格局. 生态学报, 29(5): 2211–2218. |
| [] | 巨天珍, 郝青, 葛建团, 等. 2005. 甘肃小陇山锐齿栎林空间分布格局分析. 林业资源管理(4): 27–30, 44. |
| [] | 康永祥, 岳军伟, 张巧明. 2007. 黄龙山林区辽东栎群落类型划分及其生物多样性研究. 西北林学院学报, 22(3): 7–10. |
| [] | 李荣, 张文辉, 何景峰, 等. 2011a. 不同间伐强度对辽东栎林群落稳定性的影响. 应用生态学报, 22(1): 14–20. |
| [] | 李荣, 张文辉, 何景峰, 等. 2011b. 不同间伐措施对辽东栎幼苗自然更新及生长状况的影响. 西北农林科技大学学报:自然科学版, 39(1): 52–60, 68. |
| [] | 李荣, 何景峰, 张文辉, 等. 2011c. 近自然经营间伐对辽东栎林植物组成及林木更新的影响. 西北农林科技大学学报:自然科学版, 39(8): 83–91. |
| [] | 李宗峰, 李旭光, 王永健, 等. 2007. 不同恢复群落中辽东栎种群空间格局分析. 广西植物, 27(4): 576–580. |
| [] | 缪宁, 史作民, 冯秋红, 等. 2008. 川西亚高山岷江冷杉种群的空间格局分析. 林业科学, 44(12): 1–6. DOI:10.3321/j.issn:1001-7488.2008.12.001 |
| [] | 卢彦昌, 张文辉, 陆元昌. 2006. 黄龙山林区不同培育措施对辽东栎种群结构与动态的影响. 西北植物学报, 26(7): 1407–1413. |
| [] | 王蕾, 张春雨, 赵秀海. 2009. 长白山阔叶红松林的空间分布格局. 林业科学, 45(5): 54–59. DOI:10.11707/j.1001-7488.20090508 |
| [] | 王巍, 刘灿然, 马克平, 等. 1999. 东灵山两个落叶阔叶林中辽东栎种群结构和动态. 植物学报, 41(4): 425–432. |
| [] | 王巍, 李庆康, 马克平. 2000. 东灵山地区辽东栎幼苗的建立和空间分布. 植物生态学报, 24(5): 595–600. |
| [] | 王成磊, 张文辉, 何景峰. 2011. 黄土高原丘陵沟壑区天然沙棘种群空间分布格局研究. 西北植物学报, 31(1): 152–158. |
| [] | 魏瑞, 王孝安, 郭华. 2009. 黄土高原马栏林区辽东栎的种子产量. 应用与环境生物, 15(1): 16–20. |
| [] | 伊力塔, 韩海荣, 程小琴, 等. 2008. 灵空山林区辽东栎(Quercus liaotungensis)种群空间分布格局. 生态学报, 28(7): 3254–3260. |
| [] | 张健, 郝占庆, 宋波, 等. 2007. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性. 应用生态学报, 18(8): 1681–1687. |
| [] | 张文辉, 卢志军, 李景侠, 等. 2002. 陕西不同林区栓皮栎种群空间分布格局及动态的比较研究. 西北植物学报, 22(3): 476–483. |
| [] | 张赟, 张春雨, 赵秀海, 等. 2008. 长白山次生林乔木树种空间分布格局. 生态学杂志, 27(10): 1639–1646. |
| [] | 张知彬. 2001. 埋藏和环境因子对辽东栎(Quercus liaotungensis Koidz)种子更新的影响. 生态学报, 21(3): 374–384. |
| [] | 赵则海, 杨逢建, 丛沛桐, 等. 2002. 东灵山辽东栎林木本植物多样性的研究. 植物研究, 22(4): 439–443. |
| [] | Dray S, Legendre P, Peres-Neto P R. 2006. Spatial modeling: a comprehensive framework for principal coordinate analysis of neighbor matrices (PCNM). Ecological Modelling, 196(3/4): 483–493. |
| [] | Haase. 2001. Can isotropy vs. anisotropy in the spatial association of plant species reveal physical vs. biotic facilitation. Journal of Vegetation Science, 12(1): 127–136. |
| [] | Hou J H, Mi X C, Liu C R, et al. 2004. Spatial patterns and associations in a Quercus-Betula forest in northern China. Journal of Vegetation Science, 15(3): 407–414. |
| [] | Li H J, Zhang Z B. 2003. Effect of rodents on acorn dispersal and survival of the Liaodong oak (Quercus liaotungensis Koida). Forest Ecology and Management, 176(1/3): 387–396. |
| [] | Masaki T, Ota T, Sugita H, et al. 2004. Structure and dynamics of tree populations within unsuccessful conifer plantations near the Shirakami Mountains, a snowy region of Japan. Forest Ecology and Management, 194(1/3): 389–401. |
| [] | Olsthoorn A F M, Bartelink H H, Gardiner J J, et al. 1999. Management of mixed-species forest: Silviculture and economics. DLO Institute for Forestry and Nature Research: 381–389. |
| [] | Parrotta J A, Knowles O H, Wunderle J M. 1997. Development of floristic diversity in 10-year-old restoration forests on a bauxite mined site in Amazonia. Forest Ecology and Management, 99(1/2): 21–42. |
| [] | Rigg L. 2005. Disturbance processes and spatial patterns of two emergent conifers in New Caledonia. Austral Ecology, 30(4): 363–373. DOI:10.1111/aec.2005.30.issue-4 |
| [] | Toriola D, Chereyre P, Buttler A. 1998. Distribution of primary forest plant species in a 19-year-old secondary forest in French Guiana. Journal of Tropical Ecology, 14(3): 323–340. DOI:10.1017/S026646749800025X |